

Mechanisms of hybrid oligomer formation in the pathogenesis of combined Alzheimer's and Parkinson's diseases
- PMID: 18769546
- PMCID: PMC2519786
- DOI: 10.1371/journal.pone.0003135
Mechanisms of hybrid oligomer formation in the pathogenesis of combined Alzheimer's and Parkinson's diseases
Retraction in
-
Retraction: Mechanisms of hybrid oligomer formation in the pathogenesis of combined Alzheimer's and Parkinson's diseases.PLoS One. 2025 Mar 18;20(3):e0321152. doi: 10.1371/journal.pone.0321152. eCollection 2025. PLoS One. 2025. PMID: 40100849 Free PMC article. No abstract available.
Abstract
Background: Misfolding and pathological aggregation of neuronal proteins has been proposed to play a critical role in the pathogenesis of neurodegenerative disorders. Alzheimer's disease (AD) and Parkinson's disease (PD) are frequent neurodegenerative diseases of the aging population. While progressive accumulation of amyloid beta protein (Abeta) oligomers has been identified as one of the central toxic events in AD, accumulation of alpha-synuclein (alpha-syn) resulting in the formation of oligomers and protofibrils has been linked to PD and Lewy body Disease (LBD). We have recently shown that Abeta promotes alpha-syn aggregation and toxic conversion in vivo, suggesting that abnormal interactions between misfolded proteins might contribute to disease pathogenesis. However the molecular characteristics and consequences of these interactions are not completely clear.
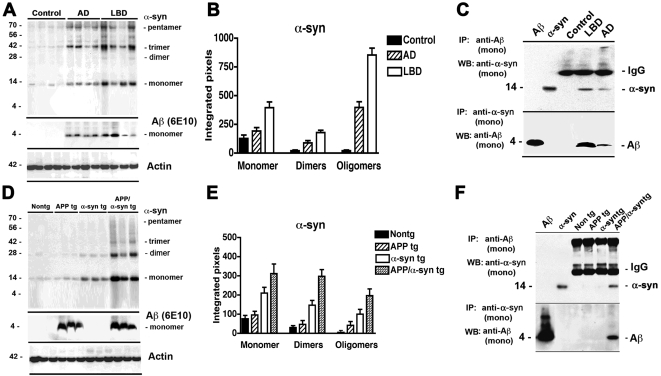
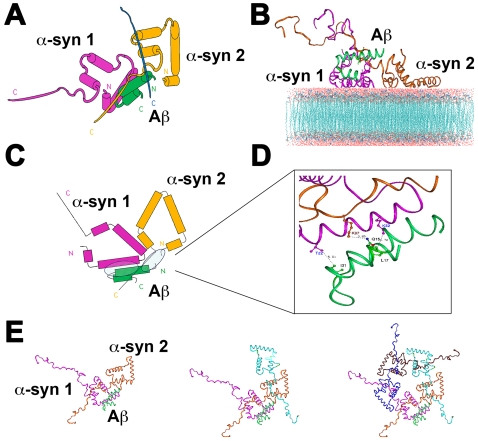
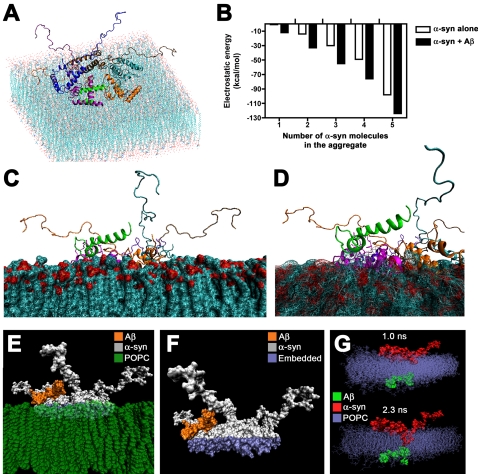
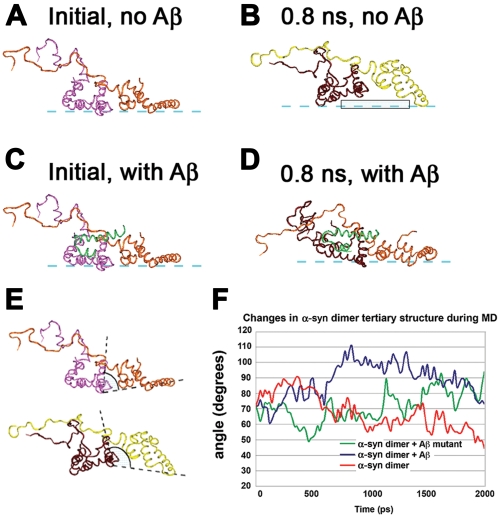
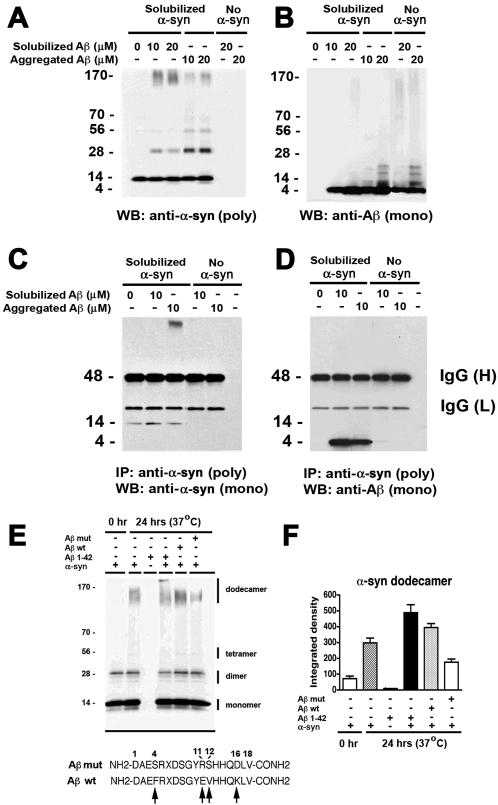
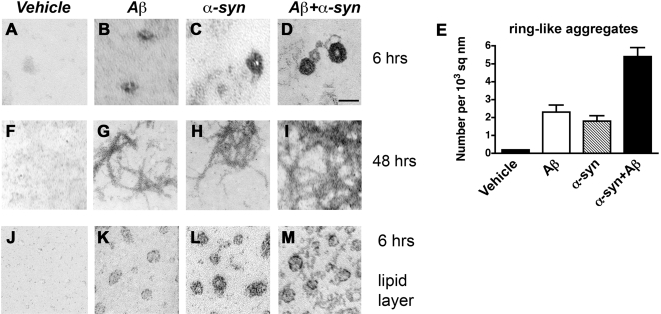
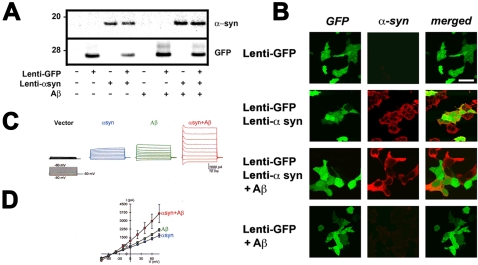
Methodology/principal findings: In order to understand the molecular mechanisms involved in potential Abeta/alpha-syn interactions, immunoblot, molecular modeling, and in vitro studies with alpha-syn and Abeta were performed. We showed in vivo in the brains of patients with AD/PD and in transgenic mice, Abeta and alpha-synuclein co-immunoprecipitate and form complexes. Molecular modeling and simulations showed that Abeta binds alpha-syn monomers, homodimers, and trimers, forming hybrid ring-like pentamers. Interactions occurred between the N-terminus of Abeta and the N-terminus and C-terminus of alpha-syn. Interacting alpha-syn and Abeta dimers that dock on the membrane incorporated additional alpha-syn molecules, leading to the formation of more stable pentamers and hexamers that adopt a ring-like structure. Consistent with the simulations, under in vitro cell-free conditions, Abeta interacted with alpha-syn, forming hybrid pore-like oligomers. Moreover, cells expressing alpha-syn and treated with Abeta displayed increased current amplitudes and calcium influx consistent with the formation of cation channels.
Conclusion/significance: These results support the contention that Abeta directly interacts with alpha-syn and stabilized the formation of hybrid nanopores that alter neuronal activity and might contribute to the mechanisms of neurodegeneration in AD and PD. The broader implications of such hybrid interactions might be important to the pathogenesis of other disorders of protein misfolding.
Conflict of interest statement
Figures
References
-
- Lashuel HA, Hartley DM, Petre BM, Wall JS, Simon MN, et al. Mixtures of wild-type and a pathogenic (E22G) form of Abeta40 in vitro accumulate protofibrils, including amyloid pores. J Mol Biol. 2003;332:795–808. - PubMed
-
- Klein WL. ADDLs & protofibrils–the missing links? Neurobiol Aging. 2002;23:231–235. - PubMed
-
- Klein WL, Krafft GA, Finch CE. Targeting small Abeta oligomers: the solution to an Alzheimer's disease conundrum? Trends Neurosci. 2001;24:219–224. - PubMed
-
- Tsigelny IF, Bar-On P, Sharikov Y, Crews L, Hashimoto M, et al. Dynamics of alpha-synuclein aggregation and inhibition of pore-like oligomer development by beta-synuclein. Febs J. 2007;274:1862–1877. - PubMed
-
- Walsh DM, Klyubin I, Fadeeva JV, Rowan MJ, Selkoe DJ. Amyloid-beta oligomers: their production, toxicity and therapeutic inhibition. Biochem Soc Trans. 2002;30:552–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
