

XIAP regulates cytosol-specific innate immunity to Listeria infection
- PMID: 18769721
- PMCID: PMC2516935
- DOI: 10.1371/journal.ppat.1000142
XIAP regulates cytosol-specific innate immunity to Listeria infection
Abstract
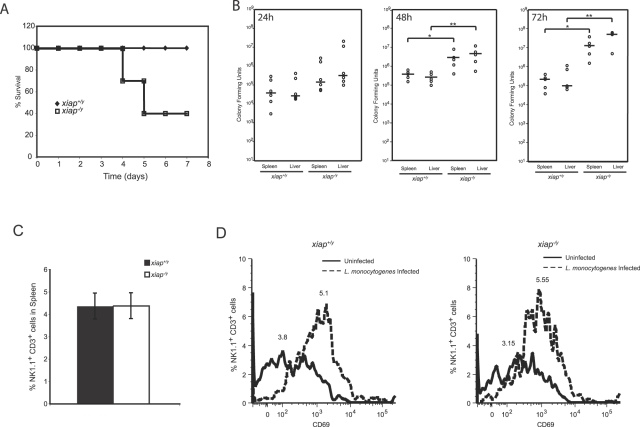
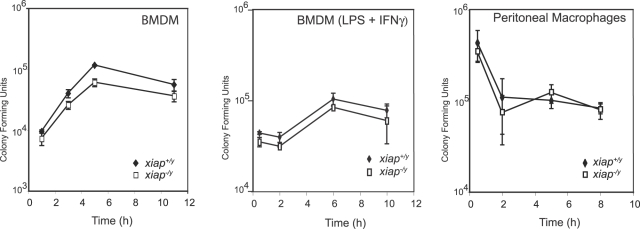
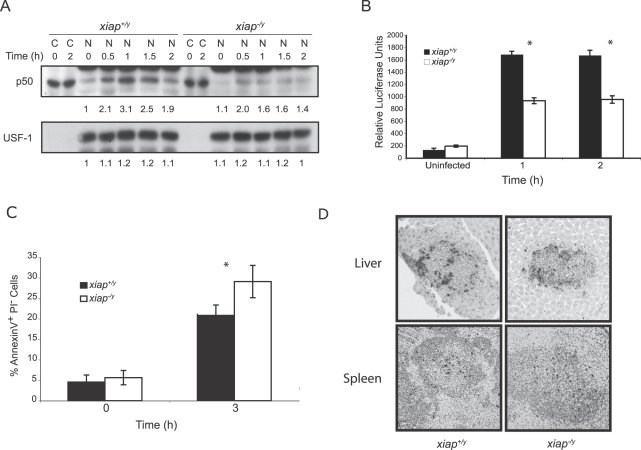
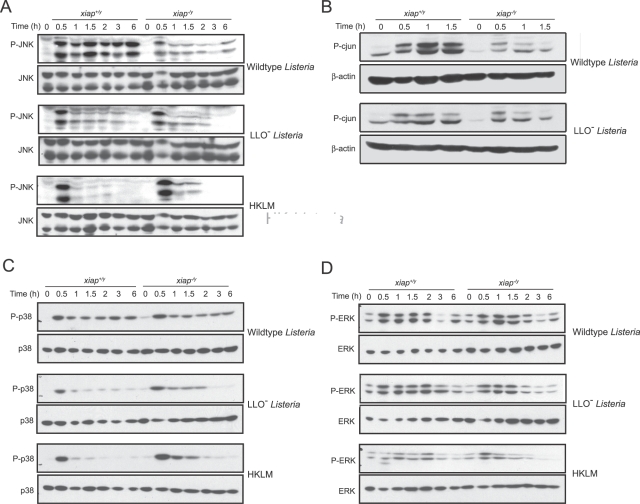
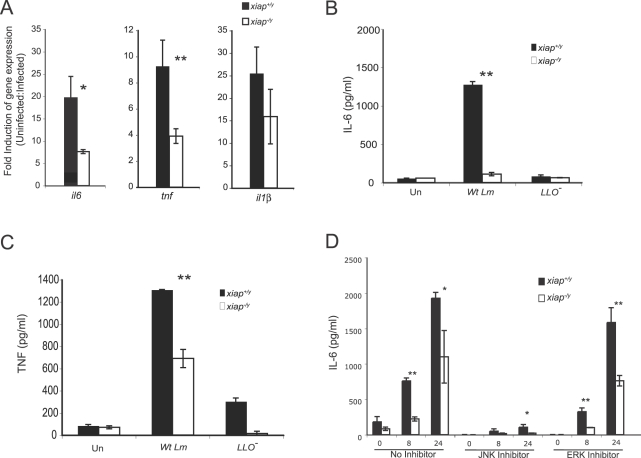
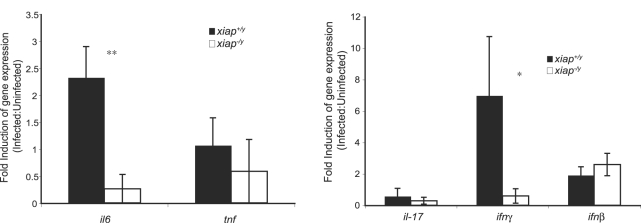
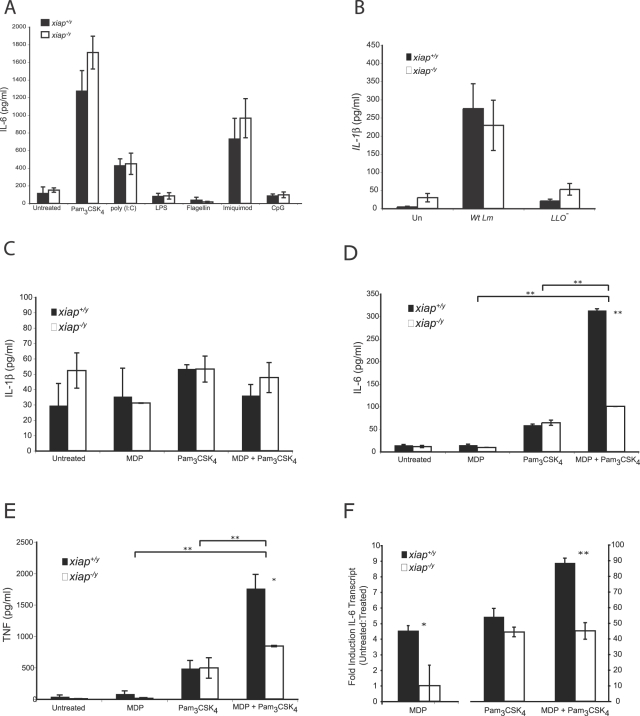
The inhibitor of apoptosis protein (IAP) family has been implicated in immune regulation, but the mechanisms by which IAP proteins contribute to immunity are incompletely understood. We show here that X-linked IAP (XIAP) is required for innate immune control of Listeria monocytogenes infection. Mice deficient in XIAP had a higher bacterial burden 48 h after infection than wild-type littermates, and exhibited substantially decreased survival. XIAP enhanced NF-kappaB activation upon L. monocytogenes infection of activated macrophages, and prolonged phosphorylation of Jun N-terminal kinase (JNK) specifically in response to cytosolic bacteria. Additionally, XIAP promoted maximal production of pro-inflammatory cytokines upon bacterial infection in vitro or in vivo, or in response to combined treatment with NOD2 and TLR2 ligands. Together, our data suggest that XIAP regulates innate immune responses to L. monocytogenes infection by potentiating synergy between Toll-like receptors (TLRs) and Nod-like receptors (NLRs) through activation of JNK- and NF-kappaB-dependent signaling.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The cytosolic sensors Nod1 and Nod2 are critical for bacterial recognition and host defense after exposure to Toll-like receptor ligands.Immunity. 2008 Feb;28(2):246-57. doi: 10.1016/j.immuni.2007.12.012. Epub 2008 Feb 7. Immunity. 2008. PMID: 18261938
-
TLR2 and RIP2 pathways mediate autophagy of Listeria monocytogenes via extracellular signal-regulated kinase (ERK) activation.J Biol Chem. 2011 Dec 16;286(50):42981-91. doi: 10.1074/jbc.M111.310599. Epub 2011 Oct 27. J Biol Chem. 2011. PMID: 22033934 Free PMC article.
-
RICK/Rip2/CARDIAK mediates signalling for receptors of the innate and adaptive immune systems.Nature. 2002 Mar 14;416(6877):194-9. doi: 10.1038/416194a. Nature. 2002. PMID: 11894098
-
Innate immune pathways triggered by Listeria monocytogenes and their role in the induction of cell-mediated immunity.Adv Immunol. 2012;113:135-56. doi: 10.1016/B978-0-12-394590-7.00002-6. Adv Immunol. 2012. PMID: 22244582 Review.
-
Listeria monocytogenes infection in the face of innate immunity.Cell Microbiol. 2009 May;11(5):703-9. doi: 10.1111/j.1462-5822.2009.01294.x. Epub 2009 Feb 2. Cell Microbiol. 2009. PMID: 19191786 Review.
Cited by
-
A regulatory region on RIPK2 is required for XIAP binding and NOD signaling activity.EMBO Rep. 2020 Nov 5;21(11):e50400. doi: 10.15252/embr.202050400. Epub 2020 Sep 21. EMBO Rep. 2020. PMID: 32954645 Free PMC article.
-
LIMP-2 links late phagosomal trafficking with the onset of the innate immune response to Listeria monocytogenes: a role in macrophage activation.J Biol Chem. 2011 Feb 4;286(5):3332-41. doi: 10.1074/jbc.M110.146761. Epub 2010 Dec 1. J Biol Chem. 2011. PMID: 21123180 Free PMC article.
-
XIAP promotes the expansion and limits the contraction of CD8 T cell response through cell extrinsic and intrinsic mechanisms respectively.PLoS Pathog. 2023 Jun 22;19(6):e1011455. doi: 10.1371/journal.ppat.1011455. eCollection 2023 Jun. PLoS Pathog. 2023. PMID: 37347786 Free PMC article.
-
S100A4 contributes to colitis development by increasing the adherence of Citrobacter rodentium in intestinal epithelial cells.Sci Rep. 2017 Sep 21;7(1):12099. doi: 10.1038/s41598-017-12256-z. Sci Rep. 2017. PMID: 28935867 Free PMC article.
-
IAPs, TNF, inflammation and Jürg Tschopp; a personal perspective.Cell Death Differ. 2012 Jan;19(1):1-4. doi: 10.1038/cdd.2011.166. Cell Death Differ. 2012. PMID: 22158430 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
