

Functional and structural stability of the epidermal growth factor receptor in detergent micelles and phospholipid nanodiscs
- PMID: 18771282
- PMCID: PMC2658769
- DOI: 10.1021/bi801006s
Functional and structural stability of the epidermal growth factor receptor in detergent micelles and phospholipid nanodiscs
Abstract
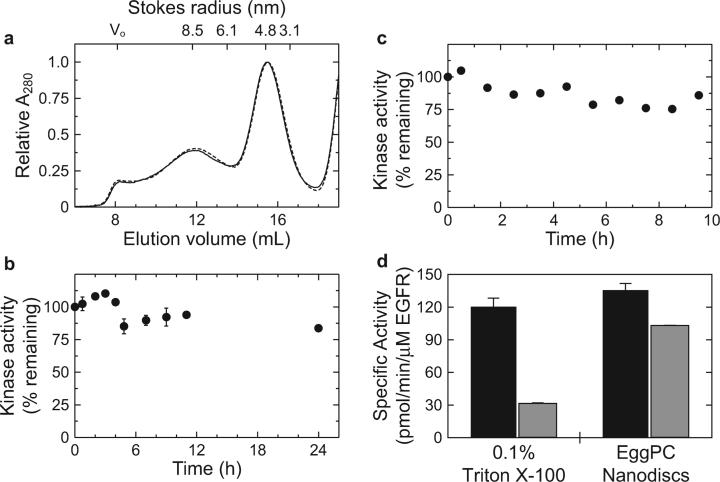
Cellular signaling mediated by the epidermal growth factor receptor (EGFR or ErbB) family of receptor tyrosine kinases plays an important role in regulating normal and oncogenic cellular physiology. While structures of isolated EGFR extracellular domains and intracellular protein tyrosine kinase domains have suggested mechanisms for growth factor-mediated receptor dimerization and allosteric kinase domain activation, understanding how the transmembrane and juxtamembrane domains contribute to transmembrane signaling requires structural studies on intact receptor molecules. In this report, recombinant EGFR constructs containing the extracellular, transmembrane, juxtamembrane, and kinase domains are overexpressed and purified from human embryonic kidney 293 cell cultures. The oligomerization state, overall structure, and functional stability of the purified EGF-bound receptor are characterized in detergent micelles and phospholipid bilayers. In the presence of EGF, catalytically active EGFR dimers can be isolated by gel filtration in dodecyl maltoside. Visualization of the dimeric species by negative stain electron microscopy and single particle averaging reveals an overall structure of the extracellular domain that is similar to previously published crystal structures and is consistent with the C-termini of domain IV being juxtaposed against one another as they enter the transmembrane domain. Although detergent-soluble preparations of EGFR are stable as dimers in the presence of EGF, they exhibit differential functional stability in Triton X-100 versus dodecyl maltoside. Furthermore, the kinase activity can be significantly stabilized by reconstituting purified EGF-bound EGFR dimers in phospholipid nanodiscs or vesicles, suggesting that the environment around the hydrophobic transmembrane and amphipathic juxtamembrane domains is important for stabilizing the tyrosine kinase activity in vitro.
Figures





Similar articles
-
Mechanisms for kinase-mediated dimerization of the epidermal growth factor receptor.J Biol Chem. 2012 Nov 2;287(45):38244-53. doi: 10.1074/jbc.M112.414391. Epub 2012 Sep 17. J Biol Chem. 2012. PMID: 22988250 Free PMC article.
-
Architecture and membrane interactions of the EGF receptor.Cell. 2013 Jan 31;152(3):557-69. doi: 10.1016/j.cell.2012.12.030. Cell. 2013. PMID: 23374350 Free PMC article.
-
Simultaneous visualization of the extracellular and cytoplasmic domains of the epidermal growth factor receptor.Nat Struct Mol Biol. 2011 Aug 7;18(9):984-9. doi: 10.1038/nsmb.2092. Nat Struct Mol Biol. 2011. PMID: 21822280 Free PMC article.
-
Regulation of the catalytic activity of the EGF receptor.Curr Opin Struct Biol. 2011 Dec;21(6):777-84. doi: 10.1016/j.sbi.2011.07.007. Epub 2011 Aug 23. Curr Opin Struct Biol. 2011. PMID: 21868214 Free PMC article. Review.
-
The ErbB/HER family of protein-tyrosine kinases and cancer.Pharmacol Res. 2014 Jan;79:34-74. doi: 10.1016/j.phrs.2013.11.002. Epub 2013 Nov 20. Pharmacol Res. 2014. PMID: 24269963 Review.
Cited by
-
Single-Molecule Fluorescence Detection of the Epidermal Growth Factor Receptor in Membrane Discs.Biochemistry. 2019 Jan 29;58(4):286-294. doi: 10.1021/acs.biochem.8b00089. Epub 2018 Apr 6. Biochemistry. 2019. PMID: 29553754 Free PMC article.
-
Nanodiscs in Membrane Biochemistry and Biophysics.Chem Rev. 2017 Mar 22;117(6):4669-4713. doi: 10.1021/acs.chemrev.6b00690. Epub 2017 Feb 8. Chem Rev. 2017. PMID: 28177242 Free PMC article. Review.
-
More than the sum of the parts: Toward full-length receptor tyrosine kinase structures.IUBMB Life. 2019 Jun;71(6):706-720. doi: 10.1002/iub.2060. Epub 2019 May 2. IUBMB Life. 2019. PMID: 31046201 Free PMC article. Review.
-
Enhanced dimerization drives ligand-independent activity of mutant epidermal growth factor receptor in lung cancer.Mol Biol Cell. 2015 Nov 5;26(22):4087-99. doi: 10.1091/mbc.E15-05-0269. Epub 2015 Sep 2. Mol Biol Cell. 2015. PMID: 26337388 Free PMC article.
-
In vitro enzymatic characterization of near full length EGFR in activated and inhibited states.Biochemistry. 2009 Jul 21;48(28):6624-32. doi: 10.1021/bi900755n. Biochemistry. 2009. PMID: 19518076 Free PMC article.
References
-
- Yarden Y, Schlessinger J. Self-phosphorylation of epidermal growth factor receptor: evidence for a model of intermolecular allosteric activation. Biochemistry. 1987;26:1434–1442. - PubMed
-
- Yarden Y, Schlessinger J. Epidermal growth factor induces rapid, reversible aggregation of the purified epidermal growth factor receptor. Biochemistry. 1987;26:1443–1451. - PubMed
-
- Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103:211–225. - PubMed
-
- Clayton AH, Walker F, Orchard SG, Henderson C, Fuchs D, Rothacker J, Nice EC, Burgess AW. Ligand-induced dimer-tetramer transition during the activation of the cell surface epidermal growth factor receptor-A multidimensional microscopy analysis. J. Biol. Chem. 2005;280:30392–30399. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
