

Engineering ascorbate peroxidase activity into cytochrome c peroxidase
- PMID: 18771292
- PMCID: PMC2770236
- DOI: 10.1021/bi8007565
Engineering ascorbate peroxidase activity into cytochrome c peroxidase
Abstract
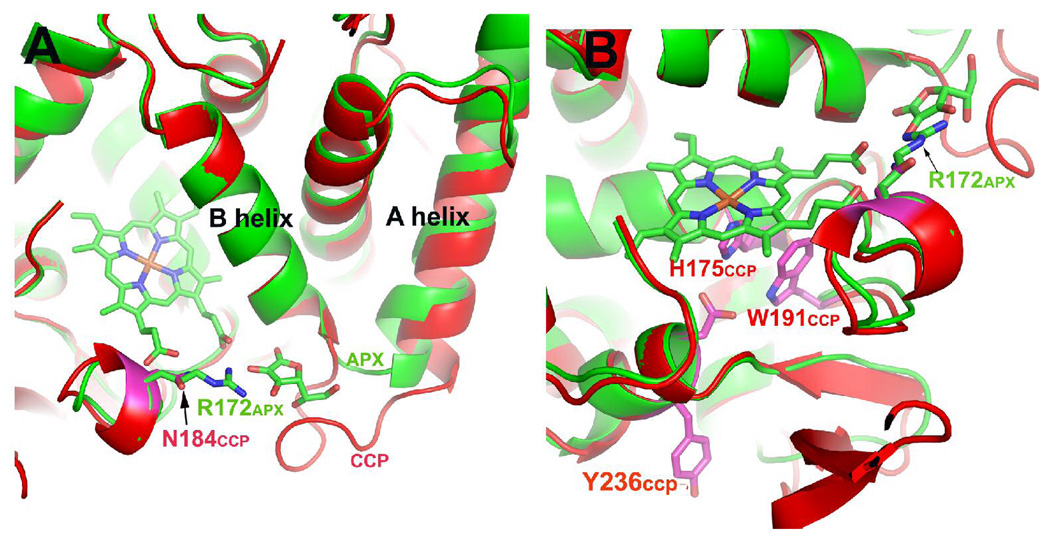
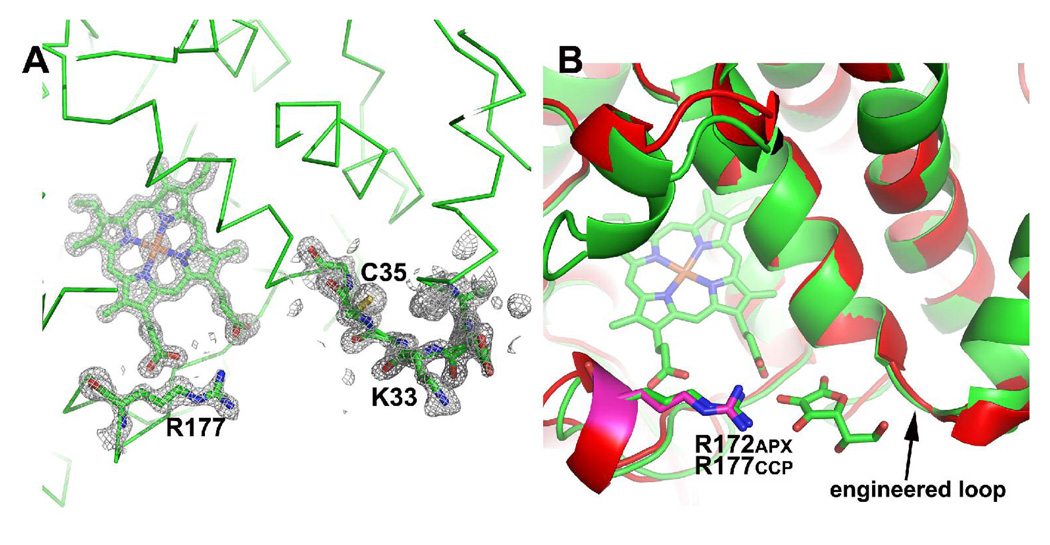
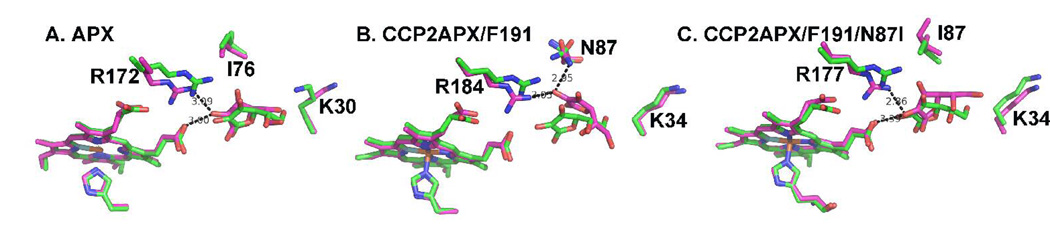
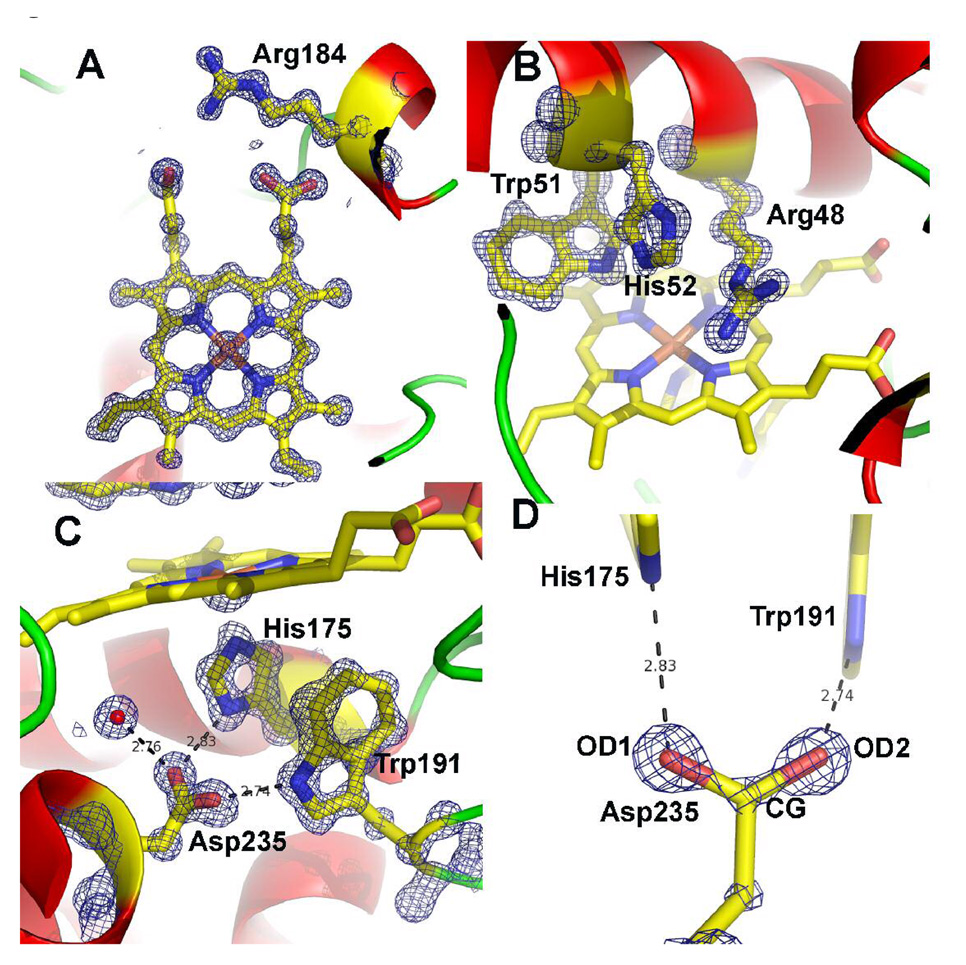
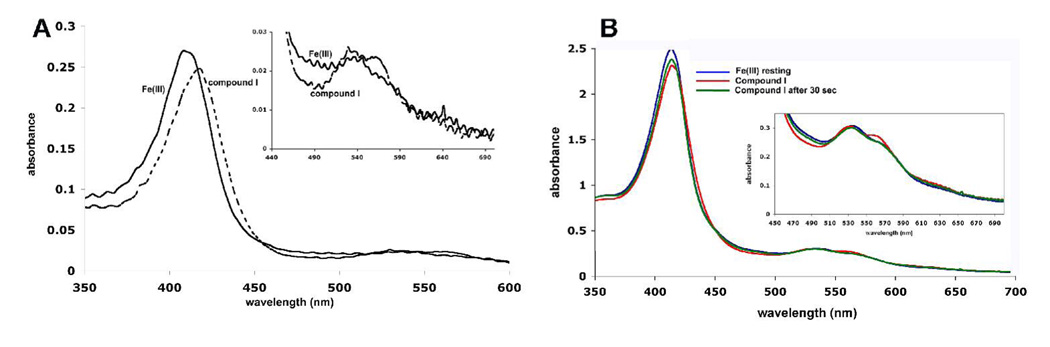
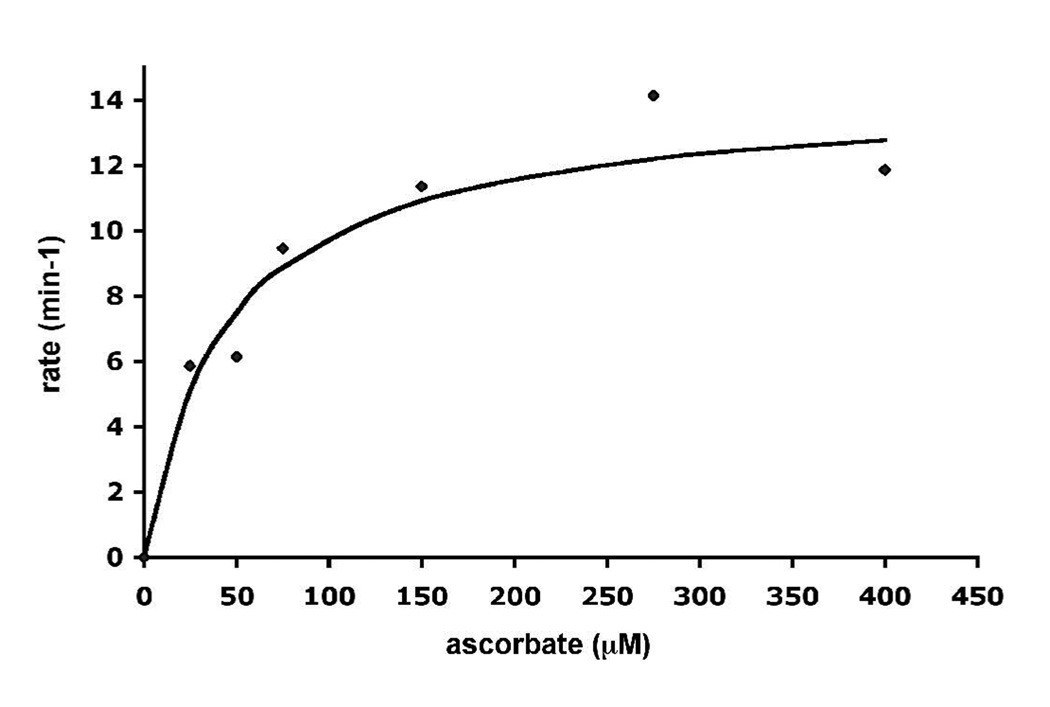
Cytochrome c peroxidase (CCP) and ascorbate peroxidase (APX) have very similar structures, and yet neither CCP nor APX exhibits each other's activities with respect to reducing substrates. APX has a unique substrate binding site near the heme propionates where ascorbate H-bonds with a surface Arg and one heme propionate (Sharp et al. (2003) Nat. Struct. Biol. 10, 303-307). The corresponding region in CCP has a much longer surface loop, and the critical Arg residue that is required for ascorbate binding in APX is Asn in CCP. In order to convert CCP into an APX, the ascorbate-binding loop and critical arginine were engineered into CCP to give the CCP2APX mutant. The mutant crystal structure shows that the engineered site is nearly identical to that found in APX. While wild-type CCP shows no APX activity, CCP2APX catalyzes the peroxidation of ascorbate at a rate of approximately 12 min (-1), indicating that the engineered ascorbate-binding loop can bind ascorbate.
Figures

Similar articles
-
Cation-induced stabilization of the engineered cation-binding loop in cytochrome c peroxidase (CcP).Biochemistry. 2002 Feb 26;41(8):2684-93. doi: 10.1021/bi011599y. Biochemistry. 2002. PMID: 11851415
-
Crystal structure of Trypanosoma cruzi heme peroxidase and characterization of its substrate specificity and compound I intermediate.J Biol Chem. 2022 Aug;298(8):102204. doi: 10.1016/j.jbc.2022.102204. Epub 2022 Jun 27. J Biol Chem. 2022. PMID: 35772495 Free PMC article.
-
The tuberculosis prodrug isoniazid bound to activating peroxidases.J Biol Chem. 2008 Mar 7;283(10):6193-200. doi: 10.1074/jbc.M707412200. Epub 2007 Dec 5. J Biol Chem. 2008. PMID: 18056997
-
Defining substrate specificity and catalytic mechanism in ascorbate peroxidase.Biochem Soc Symp. 2004;(71):27-38. doi: 10.1042/bss0710027. Biochem Soc Symp. 2004. PMID: 15777010 Review.
-
Peroxidase-catalyzed oxidation of ascorbate. Structural, spectroscopic and mechanistic correlations in ascorbate peroxidase.Subcell Biochem. 2000;35:317-49. doi: 10.1007/0-306-46828-x_10. Subcell Biochem. 2000. PMID: 11192727 Review.
Cited by
-
Designing Artificial Metalloenzymes by Tuning of the Environment beyond the Primary Coordination Sphere.Chem Rev. 2022 Jul 27;122(14):11974-12045. doi: 10.1021/acs.chemrev.2c00106. Epub 2022 Jul 11. Chem Rev. 2022. PMID: 35816578 Free PMC article. Review.
-
Ascorbate protects the diheme enzyme, MauG, against self-inflicted oxidative damage by an unusual antioxidant mechanism.Biochem J. 2017 Jul 17;474(15):2563-2572. doi: 10.1042/BCJ20170349. Biochem J. 2017. PMID: 28634178 Free PMC article.
-
A Noncanonical Tryptophan Analogue Reveals an Active Site Hydrogen Bond Controlling Ferryl Reactivity in a Heme Peroxidase.JACS Au. 2021 Jul 26;1(7):913-918. doi: 10.1021/jacsau.1c00145. Epub 2021 May 14. JACS Au. 2021. PMID: 34337604 Free PMC article.
-
Oxygen additions in serial femtosecond crystallographic protein structures.Protein Sci. 2016 Oct;25(10):1797-802. doi: 10.1002/pro.2987. Epub 2016 Jul 26. Protein Sci. 2016. PMID: 27438534 Free PMC article.
-
Crystallographic and single-crystal spectral analysis of the peroxidase ferryl intermediate.Biochemistry. 2010 Apr 13;49(14):2984-6. doi: 10.1021/bi100238r. Biochemistry. 2010. PMID: 20230048 Free PMC article.
References
-
- Sivaraja M, Goodin DB, Smith M, Hoffman BM. Identification by ENDOR of Trp 191 as the free-radical site in cytochrome c peroxidase compound ES. Science. 1989;245:738–740. - PubMed
-
- Dunford HB. In: Horseradish peroxidase: structure and kinetic properties. Everse J, Everse KE, Grisham MB, editors. Boca Raton: CRC Press; 1991. pp. 1–24.
-
- Henriksen A, Smith AT, Gajhede MY. The structures of the horseradish peroxidase C-ferulic acid complex and the ternary complex with cyanide suggest how peroxidases oxidize small phenolic substrates. J. Biol. Chem. 1999;274:35005–35011. - PubMed
-
- Pelletier H, Kraut J. Crystal structure of a complex between electron transfer partners, cytochrome c peroxidase and cytochrome c. Science. 1992;258:1748–1755. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
