

Smad-independent transforming growth factor-beta regulation of early growth response-1 and sustained expression in fibrosis: implications for scleroderma
- PMID: 18772333
- PMCID: PMC2543076
- DOI: 10.2353/ajpath.2008.080382
Smad-independent transforming growth factor-beta regulation of early growth response-1 and sustained expression in fibrosis: implications for scleroderma
Abstract
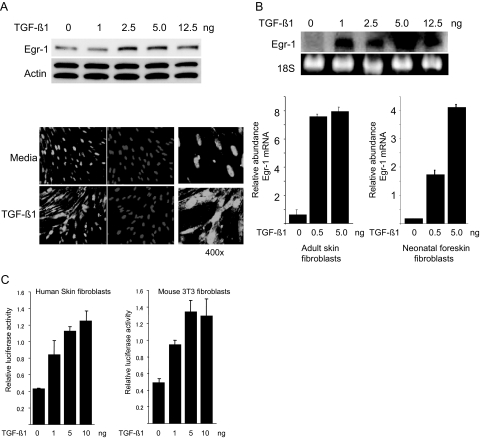
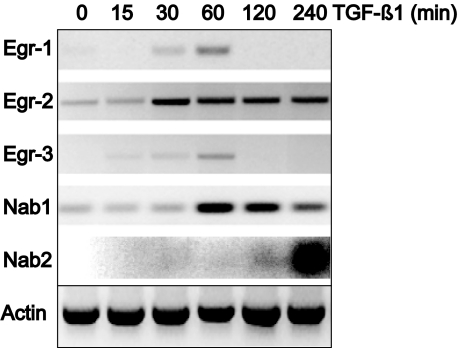
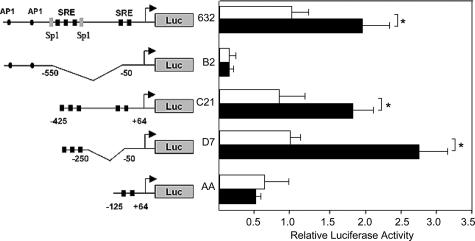
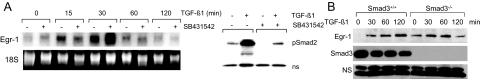
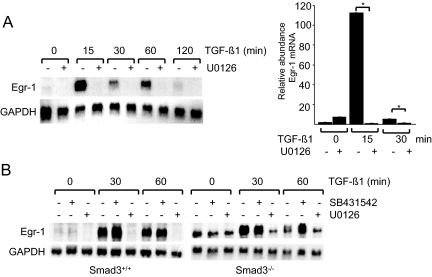
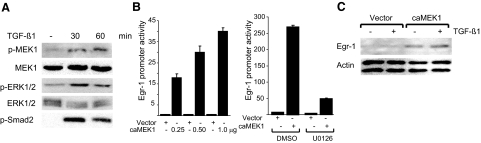
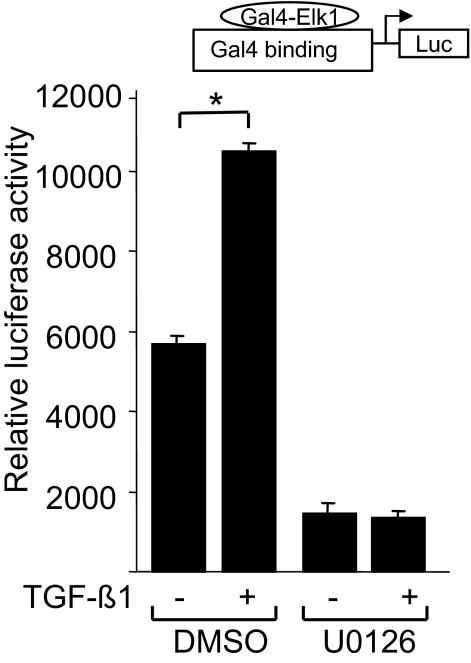
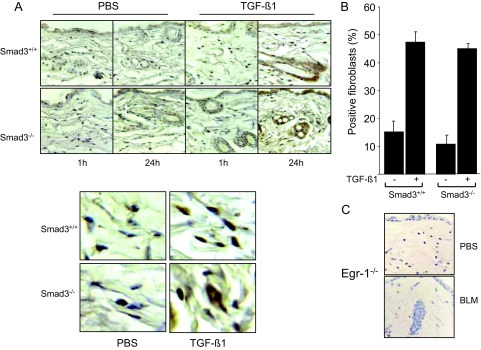
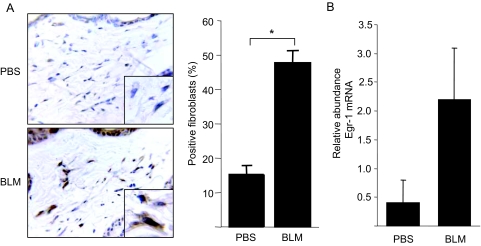
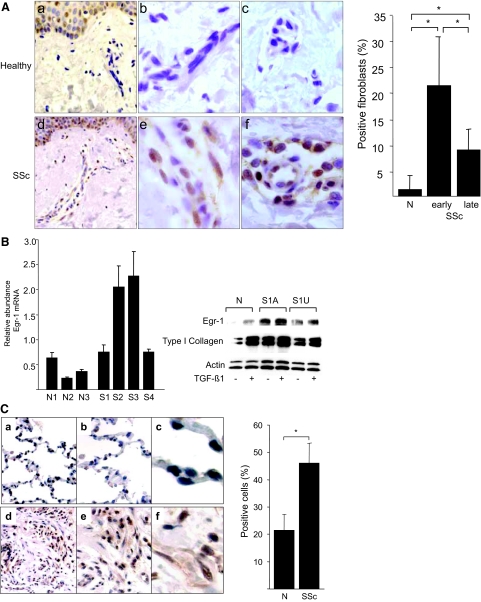
Transforming growth factor-beta (TGF-beta) plays a key role in scleroderma pathogenesis. The transcription factor early growth response-1 (Egr-1) mediates the stimulation of collagen transcription elicited by TGF-beta and is necessary for the development of pulmonary fibrosis in mice. Here, we report that TGF-beta causes a time- and dose-dependent increase in Egr-1 protein and mRNA levels and enhanced transcription of the Egr-1 gene via serum response elements in normal fibroblasts. The ability of TGF-beta to stimulate Egr-1 was preserved in Smad3-null mice and in explanted Smad3-null fibroblasts. The response was blocked by a specific mitogen-activated protein kinase kinase 1 (MEK1) inhibitor but not by an ALK5 kinase inhibitor. Furthermore, MEK1 was phosphorylated by TGF-beta, which was sufficient to drive Egr-1 transactivation. Stimulation by TGF-beta enhanced the transcriptional activity of Elk-1 via the MEK-extracellular signal-regulated kinase 1/2 pathway. Bleomycin-induced scleroderma in the mouse was accompanied by increased Egr-1 accumulation in lesional fibroblasts. Furthermore, biopsies of lesional skin and lung from patients with scleroderma showed increased Egr-1 levels, which were highest in early diffuse disease. Moreover, both Egr-1 mRNA and protein were elevated in explanted scleroderma skin fibroblasts in vitro. Together, these findings define a Smad-independent TGF-beta signal transduction mechanism that underlies the stimulation of Egr-1, demonstrate for the first time sustained Egr-1 up-regulation in fibrotic lesions and suggests that Egr-1 has a role in the induction and progression of fibrosis.
Figures
References
-
- Jimenez SA, Derk CT. Following the molecular pathways toward an understanding of the pathogenesis of systemic sclerosis. Ann Intern Med. 2004;140:37–45. - PubMed
-
- Mauviel A. Transforming growth factor-beta: a key mediator of fibrosis. Methods Mol Med. 2005;117:69–80. - PubMed
-
- Massagué J, Seoane J, Wotton D. Smad transcription factors (review). Genes Dev. 2005;19:2783–2810. - PubMed
-
- Moustakas A, Heldin CH. Non-Smad TGF-beta signals (review). J Cell Sci. 2005;118:3573–3584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
