

Loss of PIP5KIbeta demonstrates that PIP5KI isoform-specific PIP2 synthesis is required for IP3 formation
- PMID: 18772378
- PMCID: PMC2544579
- DOI: 10.1073/pnas.0804139105
Loss of PIP5KIbeta demonstrates that PIP5KI isoform-specific PIP2 synthesis is required for IP3 formation
Erratum in
- Proc Natl Acad Sci U S A. 2009 Jan 27;106(4):1291. Kanaho, Yasunori [added]
Abstract
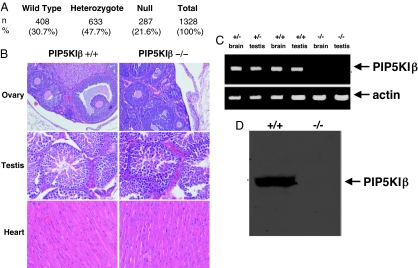
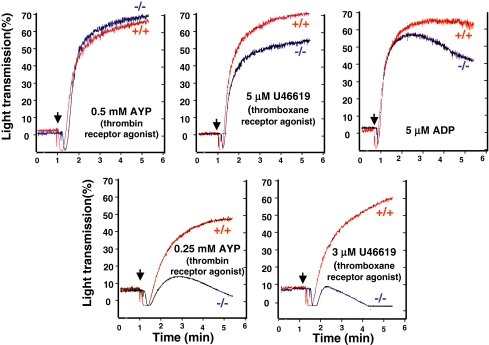
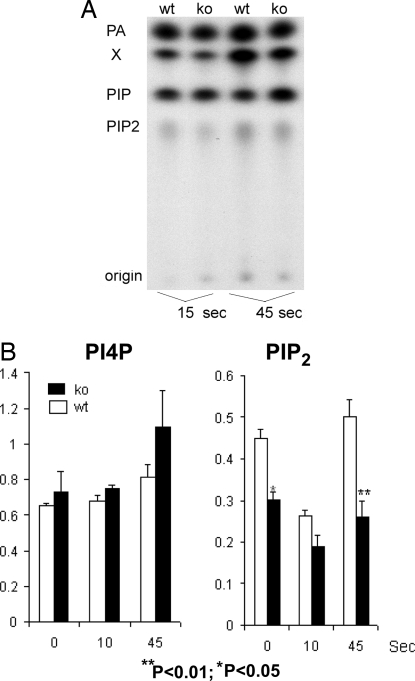
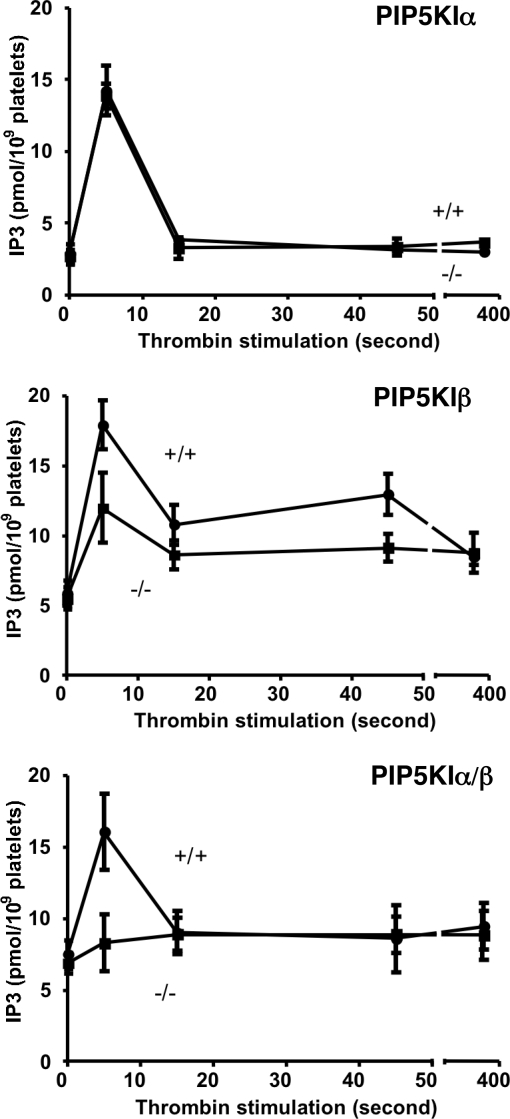
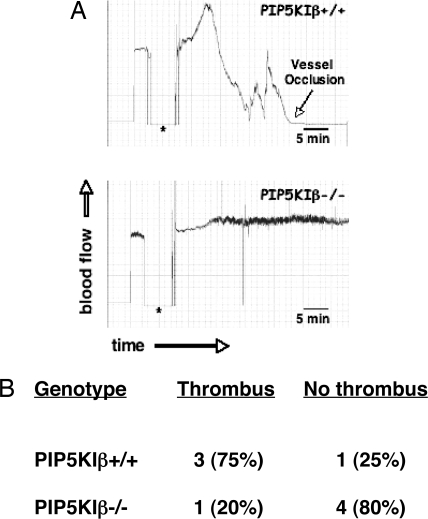
The three isoforms of PIP5KI (alpha, beta, and gamma) synthesize PI4,5P(2) (PIP(2)) by phosphorylating PI4P. Therefore, it is not clear why platelets, like all eukaryotic cells, have more than one isoform. To test the hypothesis that PIP5KI isoforms have nonoverlapping functions, we generated a murine line containing a null mutation of PIP5KIbeta and analyzed the effect on platelet signaling. PIP5KIbeta-null mice had normal platelet counts. In contrast to platelets lacking PIP5KIalpha, platelets lacking PIP5KIbeta exhibited impaired aggregation accompanied by disaggregation. Although platelets lacking PIP5KIbeta had only a moderate deficiency of PIP(2) under basal conditions, they had a striking deficiency in PIP(2) synthesis and IP(3) formation after thrombin stimulation. We have also observed that platelets lacking both PIP5KIalpha and PIP5KIbeta have a complete loss of thrombin-induced IP(3) synthesis even though they still contain PIP5KIgamma, the predominant PIP5KI isoform in platelets. These results demonstrate that PIP5KIbeta, like PIP5KIalpha, contributes to the rapid synthesis of a pool of PIP(2) that is required for second-messenger formation, whereas the pool of PIP(2) synthesized by PIP5KIgamma does not contribute to this process. Additionally, we found that PIP5KIbeta-null platelets failed to form arterial thrombi properly in vivo. Together, these data demonstrate that PIP5KIbeta is required for rapid PIP(2) synthesis, second-messenger production, and stable platelet adhesion under shear in vivo. These results also demonstrate that after stimulation of a G protein-coupled receptor, IP(3) is completely derived from a rapidly synthesized discrete pool of PIP(2) synthesized by PIP5KIalpha and PIP5KIbeta.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Gilmore AP, Burridge K. Regulation of vinculin binding to talin and actin by phosphatidylinositol 4,5-bisphosphate. Nature. 1996;381:531–535. - PubMed
-
- Furuhashi K, et al. Inositol phospholipid-induced suppression of F-actin-gelating activity of smooth muscle filamin. Biochem Biophys Res Commun. 1992;184:1261–1265. - PubMed
-
- Stephens LR, Hughes KT, Irvine RF. Pathway of phosphatidylinositol(3,4,5)-trisphosphate synthesis in activated neutrophils. Nature. 1991;351:33–39. - PubMed
-
- Ishihara H, et al. Cloning of cDNAs encoding two isoforms of 68-kDa type I phosphatidylinositol-4-phosphate 5-kinase. J Biol Chem. 1996;271:23611–23614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
