

Maternal Groucho and bHLH repressors amplify the dose-sensitive X chromosome signal in Drosophila sex determination
- PMID: 18773886
- PMCID: PMC2653429
- DOI: 10.1016/j.ydbio.2008.08.012
Maternal Groucho and bHLH repressors amplify the dose-sensitive X chromosome signal in Drosophila sex determination
Abstract
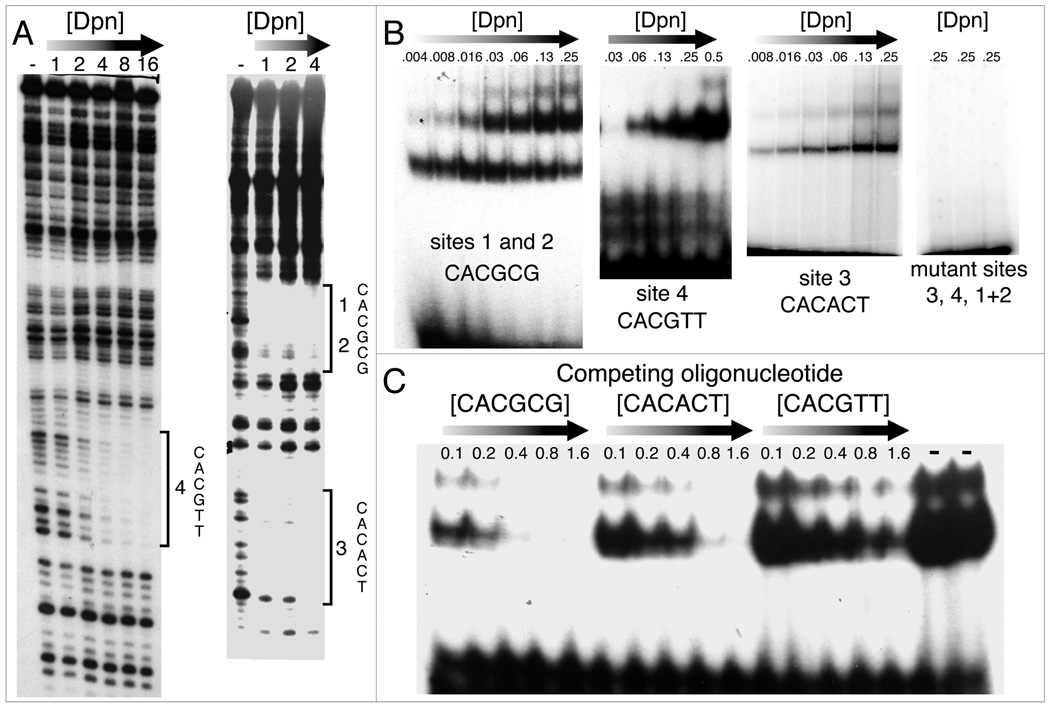
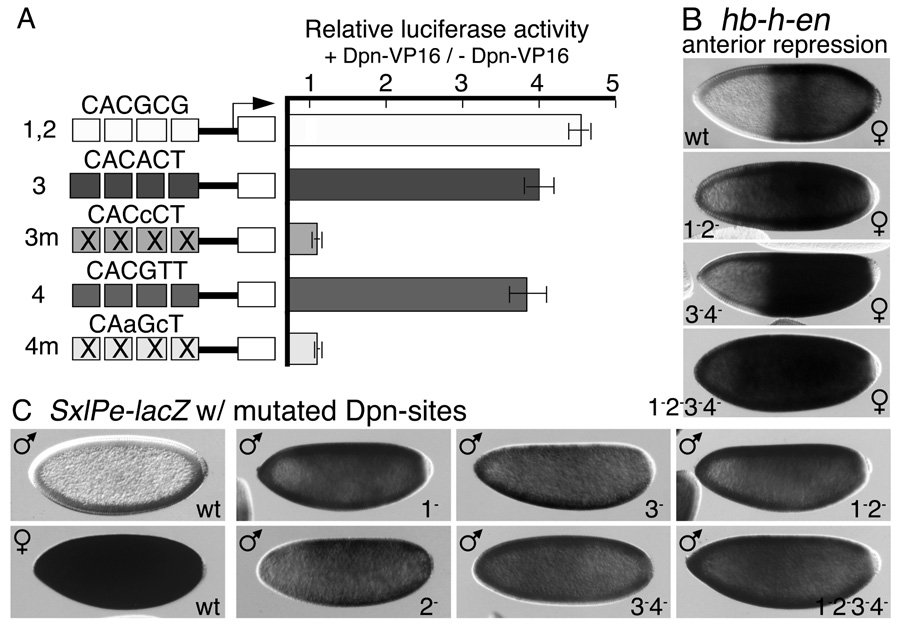
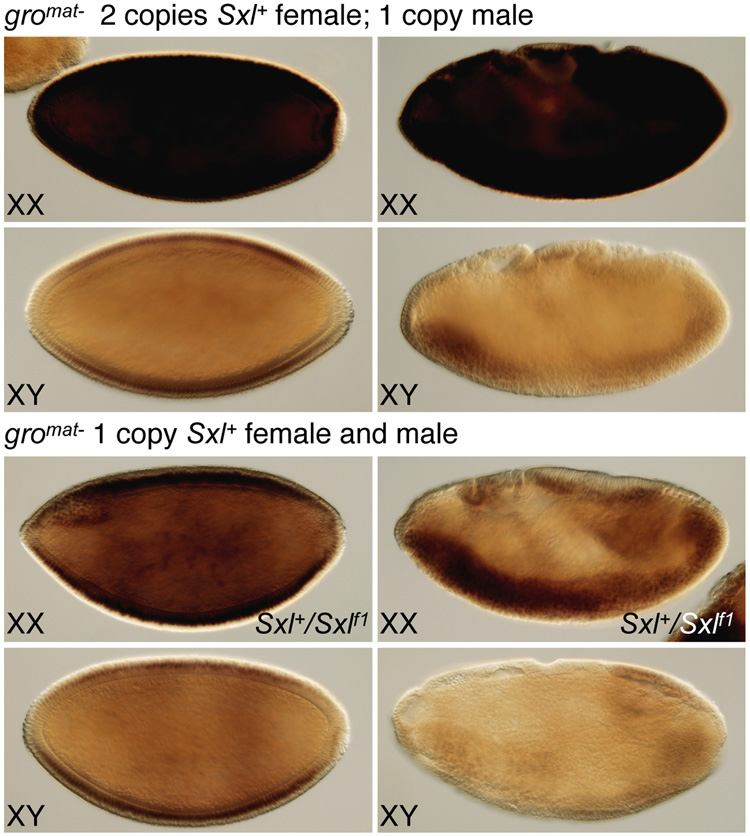
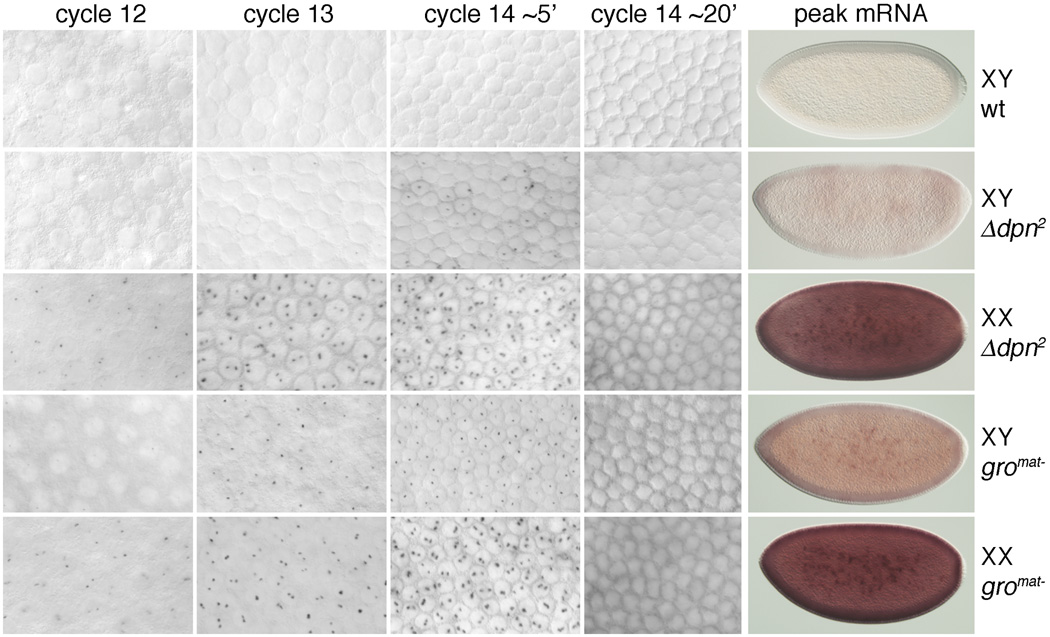
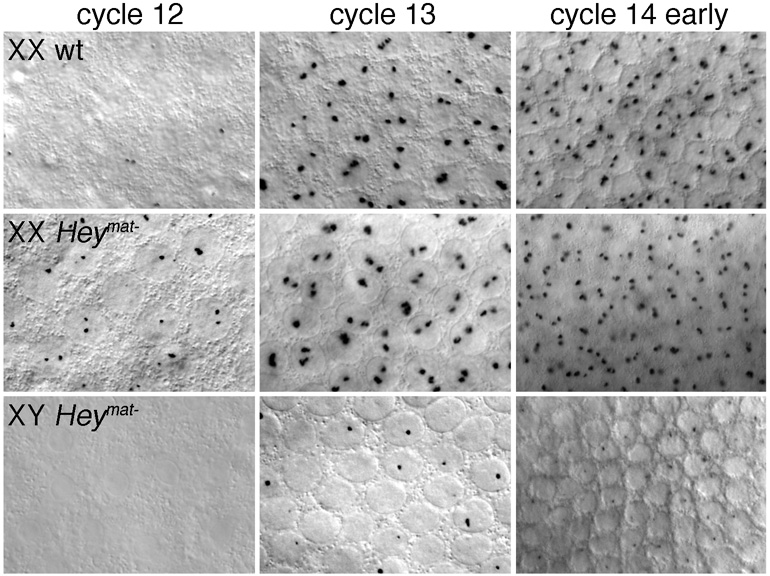
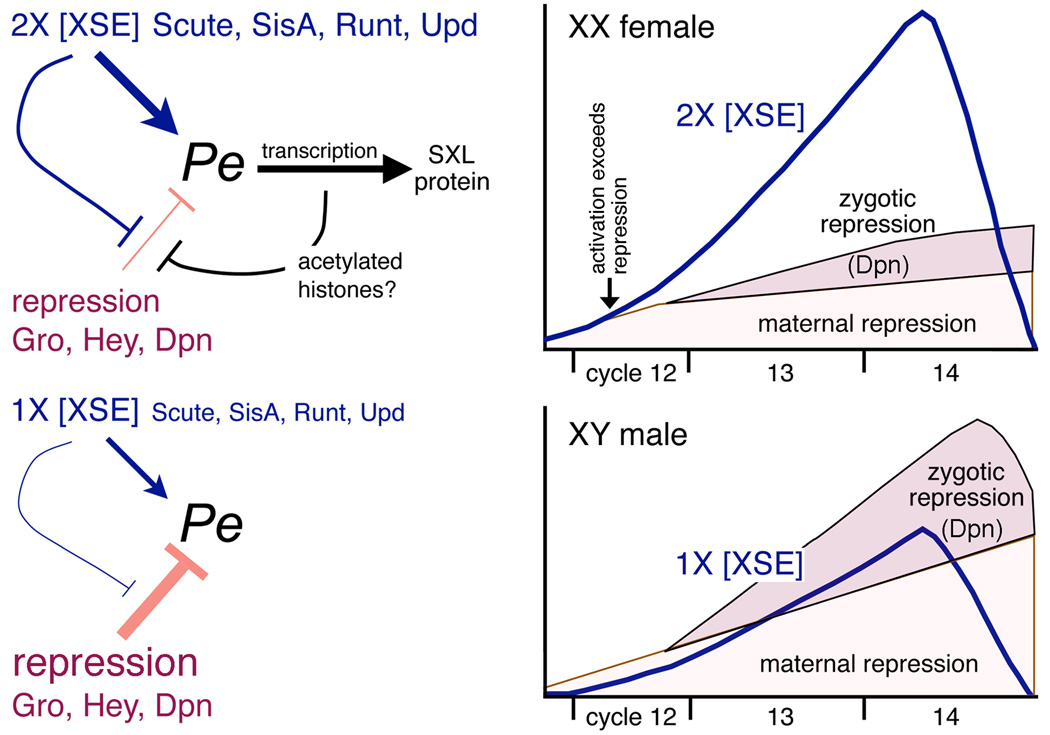
In Drosophila, XX embryos are fated to develop as females, and XY embryos as males, because the diplo-X dose of four X-linked signal element genes, XSEs, activates the Sex-lethal establishment promoter, SxlPe, whereas the haplo-X XSE dose leaves SxlPe off. The threshold response of SxlPe to XSE concentrations depends in part on the bHLH repressor, Deadpan, present in equal amounts in XX and XY embryos. We identified canonical and non-canonical DNA-binding sites for Dpn at SxlPe and found that cis-acting mutations in the Dpn-binding sites caused stronger and earlier Sxl expression than did deletion of dpn implicating other bHLH repressors in Sxl regulation. Maternal Hey encodes one such bHLH regulator but the E(spl) locus does not. Elimination of the maternal corepressor Groucho also caused strong ectopic Sxl expression in XY, and premature Sxl activation in XX embryos, but Sxl was still expressed differently in the sexes. Our findings suggest that Groucho and associated maternal and zygotic bHLH repressors define the threshold XSE concentrations needed to activate SxlPe and that they participate directly in sex signal amplification. We present a model in which the XSE signal is amplified by a feedback mechanism that interferes with Gro-mediated repression in XX, but not XY embryos.
Figures

Similar articles
-
Evidence That Runt Acts as a Counter-Repressor of Groucho During Drosophila melanogaster Primary Sex Determination.G3 (Bethesda). 2020 Jul 7;10(7):2487-2496. doi: 10.1534/g3.120.401384. G3 (Bethesda). 2020. PMID: 32457096 Free PMC article.
-
Drosophila JAK/STAT pathway reveals distinct initiation and reinforcement steps in early transcription of Sxl.Curr Biol. 2007 Apr 3;17(7):643-8. doi: 10.1016/j.cub.2007.02.038. Epub 2007 Mar 15. Curr Biol. 2007. PMID: 17363251
-
Interpretation of X chromosome dose at Sex-lethal requires non-E-box sites for the basic helix-loop-helix proteins SISB and daughterless.Mol Cell Biol. 2001 Mar;21(5):1581-92. doi: 10.1128/MCB.21.5.1581-1592.2001. Mol Cell Biol. 2001. PMID: 11238895 Free PMC article.
-
E(spl): genetic, developmental, and evolutionary aspects of a group of invertebrate Hes proteins with close ties to Notch signaling.Curr Top Dev Biol. 2014;110:217-62. doi: 10.1016/B978-0-12-405943-6.00006-3. Curr Top Dev Biol. 2014. PMID: 25248478 Review.
-
Sex determination in Drosophila: The view from the top.Fly (Austin). 2010 Jan-Mar;4(1):60-70. doi: 10.4161/fly.4.1.11277. Epub 2010 Jan 21. Fly (Austin). 2010. PMID: 20160499 Free PMC article. Review.
Cited by
-
Evidence That Runt Acts as a Counter-Repressor of Groucho During Drosophila melanogaster Primary Sex Determination.G3 (Bethesda). 2020 Jul 7;10(7):2487-2496. doi: 10.1534/g3.120.401384. G3 (Bethesda). 2020. PMID: 32457096 Free PMC article.
-
The transcription factor Hey and nuclear lamins specify and maintain cell identity.Elife. 2019 Jul 16;8:e44745. doi: 10.7554/eLife.44745. Elife. 2019. PMID: 31310235 Free PMC article.
-
Transcriptome Analysis of the Oriental Fruit Fly Bactrocera dorsalis Early Embryos.Insects. 2020 May 23;11(5):323. doi: 10.3390/insects11050323. Insects. 2020. PMID: 32456171 Free PMC article.
-
High-affinity biomolecular interactions are modulated by low-affinity binders.NPJ Syst Biol Appl. 2024 Aug 10;10(1):85. doi: 10.1038/s41540-024-00410-z. NPJ Syst Biol Appl. 2024. PMID: 39127695 Free PMC article.
-
Transcriptome analysis of sex-biased gene expression in the spotted-wing Drosophila, Drosophila suzukii (Matsumura).G3 (Bethesda). 2022 Jul 29;12(8):jkac127. doi: 10.1093/g3journal/jkac127. G3 (Bethesda). 2022. PMID: 35587603 Free PMC article.
References
-
- Ashburner M, Golic K, Hawley R. Drosophila A Laboratory Handbook. Cold Spring Harbor, New York: Cold Spring Harbor Press; 2005.
-
- Avila FW, Erickson JW. Drosophila JAK/STAT pathway reveals distinct initiation and reinforcement steps in early transcription of Sxl. Curr Biol. 2007;17:643–648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
