

The role of nucleotide cofactor binding in cooperativity and specificity of MutS recognition
- PMID: 18773911
- PMCID: PMC2666446
- DOI: 10.1016/j.jmb.2008.08.052
The role of nucleotide cofactor binding in cooperativity and specificity of MutS recognition
Abstract
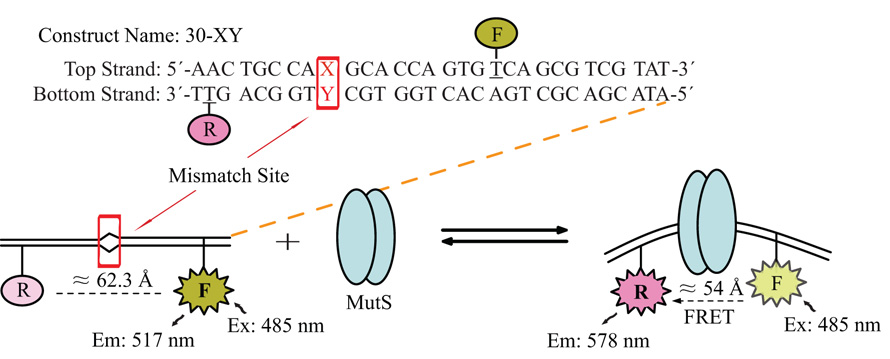
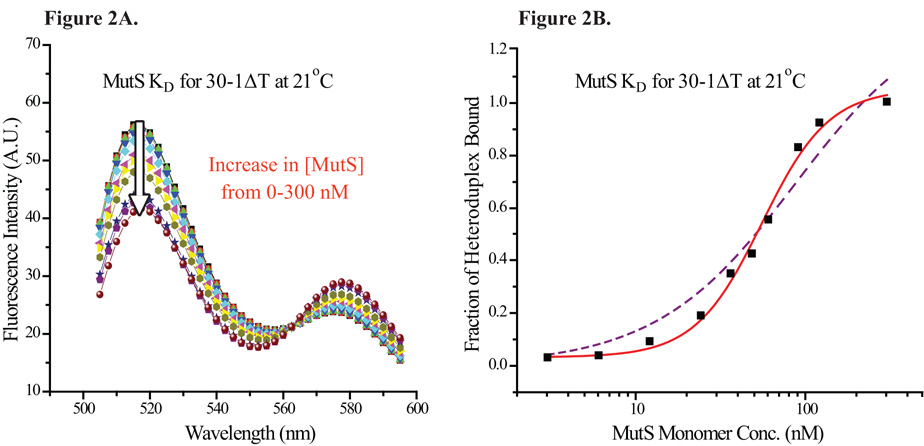
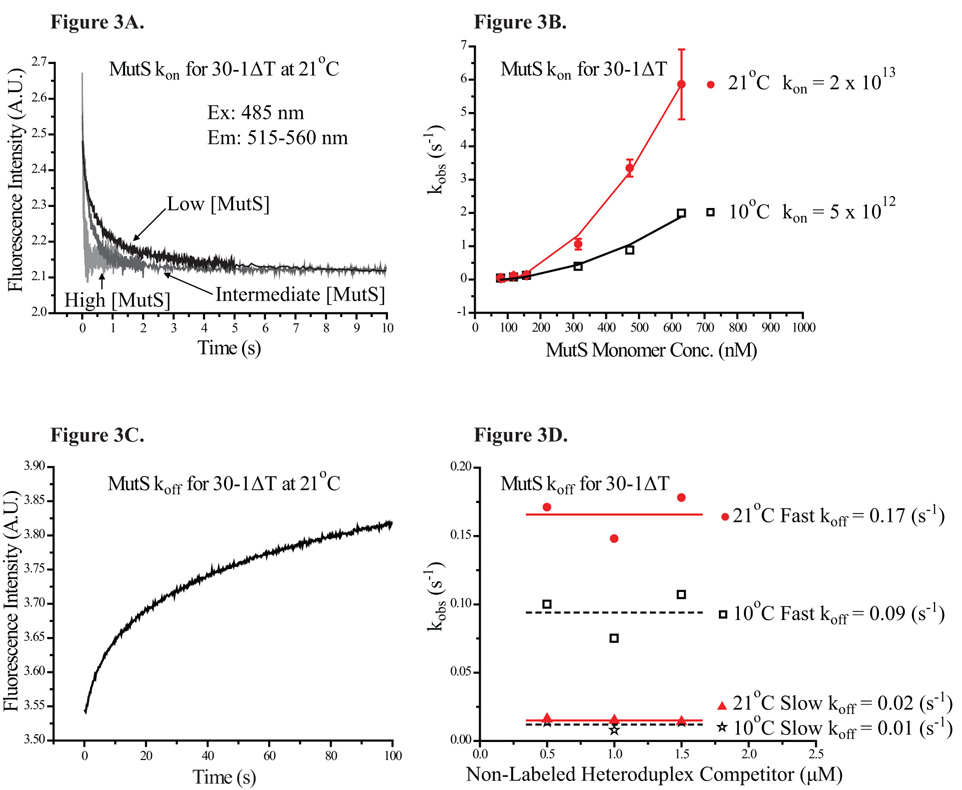
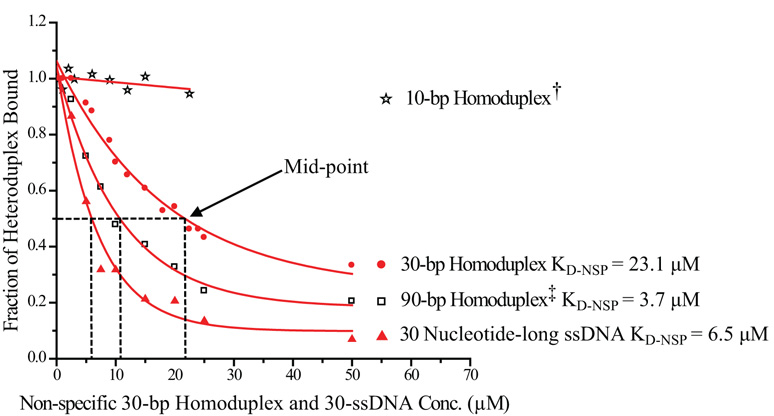
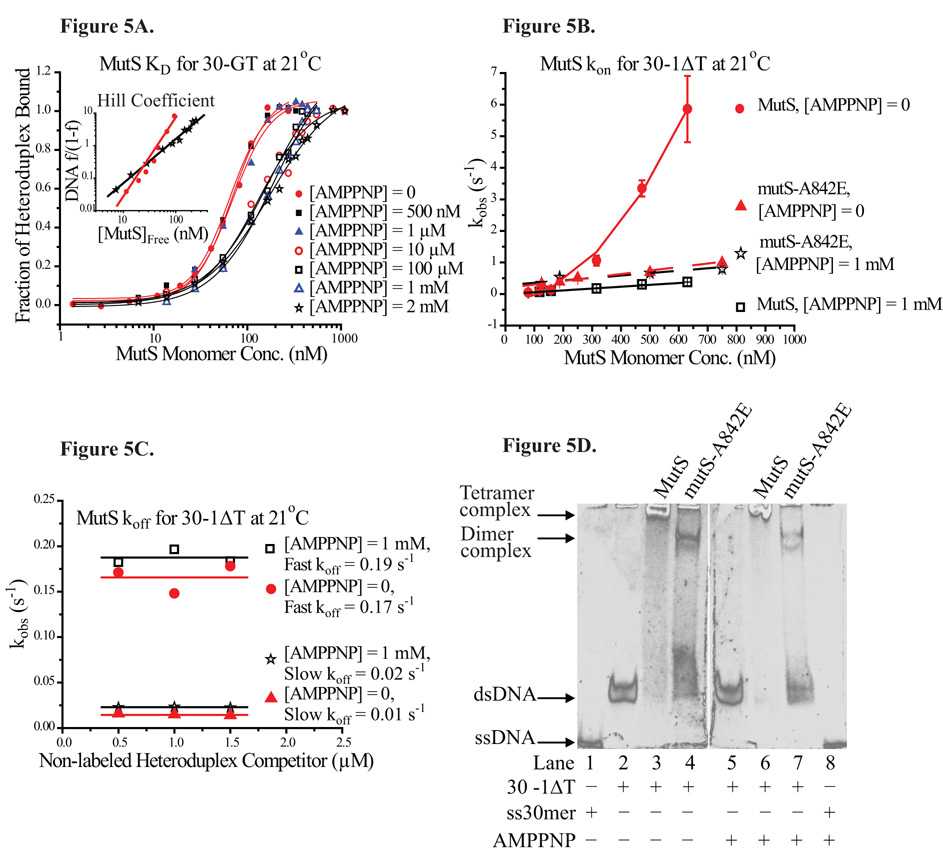
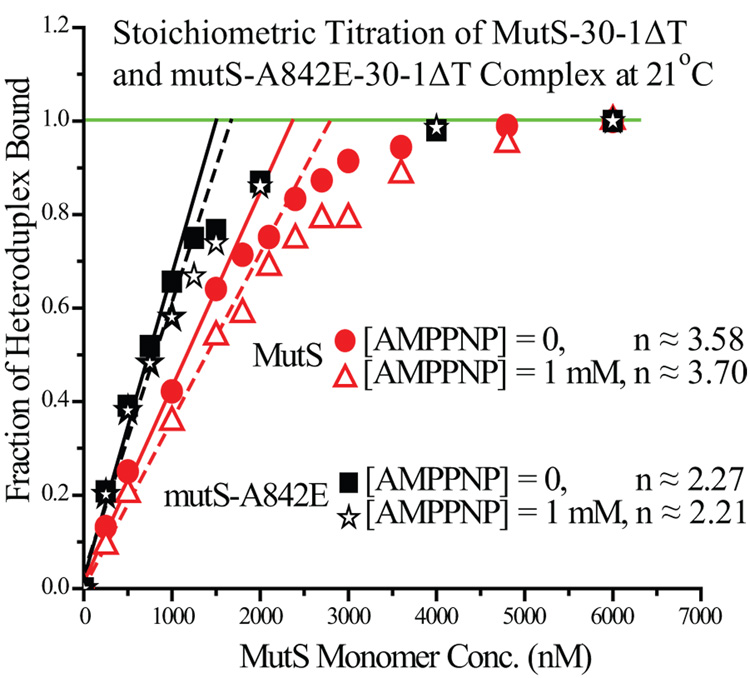
Mismatch repair (MMR) is essential for eliminating biosynthetic errors generated during replication or genetic recombination in virtually all organisms. The critical first step in Escherichia coli MMR is the specific recognition and binding of MutS to a heteroduplex, containing either a mismatch or an insertion/deletion loop of up to four nucleotides. All known MutS homologs recognize a similar broad spectrum of substrates. Binding and hydrolysis of nucleotide cofactors by the MutS-heteroduplex complex are required for downstream MMR activity, although the exact role of the nucleotide cofactors is less clear. Here, we showed that MutS bound to a 30-bp heteroduplex containing an unpaired T with a binding affinity approximately 400-fold stronger than to a 30-bp homoduplex, a much higher specificity than previously reported. The binding of nucleotide cofactors decreased both MutS specific and nonspecific binding affinity, with the latter marked by a larger drop, further increasing MutS specificity by approximately 3-fold. Kinetic studies showed that the difference in MutS K(d) for various heteroduplexes was attributable to the difference in intrinsic dissociation rate of a particular MutS-heteroduplex complex. Furthermore, the kinetic association event of MutS binding to heteroduplexes was marked by positive cooperativity. Our studies showed that the positive cooperativity in MutS binding was modulated by the binding of nucleotide cofactors. The binding of nucleotide cofactors transformed E. coli MutS tetramers, the functional unit in E. coli MMR, from a cooperative to a noncooperative binding form. Finally, we found that E. coli MutS bound to single-strand DNA with significant affinity, which could have important implication for strand discrimination in eukaryotic MMR mechanism.
Figures
Similar articles
-
Modulation of MutS ATP-dependent functional activities by DNA containing a cisplatin compound lesion (base damage and mismatch).J Mol Biol. 2007 May 25;369(1):27-40. doi: 10.1016/j.jmb.2007.02.048. Epub 2007 Feb 22. J Mol Biol. 2007. PMID: 17400248
-
Distinct MutS DNA-binding modes that are differentially modulated by ATP binding and hydrolysis.J Biol Chem. 2001 Sep 7;276(36):34339-47. doi: 10.1074/jbc.M104256200. Epub 2001 Jul 13. J Biol Chem. 2001. PMID: 11454861
-
G-quadruplex recognition activities of E. Coli MutS.BMC Mol Biol. 2012 Jul 2;13:23. doi: 10.1186/1471-2199-13-23. BMC Mol Biol. 2012. PMID: 22747774 Free PMC article.
-
DNA mismatch repair and mutation avoidance pathways.J Cell Physiol. 2002 Apr;191(1):28-41. doi: 10.1002/jcp.10077. J Cell Physiol. 2002. PMID: 11920679 Review.
-
Single-molecule views of MutS on mismatched DNA.DNA Repair (Amst). 2014 Aug;20:82-93. doi: 10.1016/j.dnarep.2014.02.014. Epub 2014 Mar 12. DNA Repair (Amst). 2014. PMID: 24629484 Free PMC article. Review.
Cited by
-
Evidence for a bind-then-bend mechanism for architectural DNA binding protein yNhp6A.Nucleic Acids Res. 2019 Apr 8;47(6):2871-2883. doi: 10.1093/nar/gkz022. Nucleic Acids Res. 2019. PMID: 30698746 Free PMC article.
-
Using stable MutS dimers and tetramers to quantitatively analyze DNA mismatch recognition and sliding clamp formation.Nucleic Acids Res. 2013 Sep;41(17):8166-81. doi: 10.1093/nar/gkt582. Epub 2013 Jul 1. Nucleic Acids Res. 2013. PMID: 23821665 Free PMC article.
-
Two-step interrogation then recognition of DNA binding site by Integration Host Factor: an architectural DNA-bending protein.Nucleic Acids Res. 2018 Feb 28;46(4):1741-1755. doi: 10.1093/nar/gkx1215. Nucleic Acids Res. 2018. PMID: 29267885 Free PMC article.
-
Identification of a mismatch-specific endonuclease in hyperthermophilic Archaea.Nucleic Acids Res. 2016 Apr 20;44(7):2977-86. doi: 10.1093/nar/gkw153. Epub 2016 Mar 21. Nucleic Acids Res. 2016. PMID: 27001046 Free PMC article.
-
Postreplicative mismatch repair.Cold Spring Harb Perspect Biol. 2013 Apr 1;5(4):a012633. doi: 10.1101/cshperspect.a012633. Cold Spring Harb Perspect Biol. 2013. PMID: 23545421 Free PMC article. Review.
References
-
- Modrich P, Lahue R. Mismatch repair in replication fidelity, genetic recombination, and cancer biology. Annu Rev Biochem. 1996;65:101–133. - PubMed
-
- Schofield MJ, Hsieh P. DNA mismatch repair: molecular mechanisms and biological function. Annu Rev Microbiol. 2003;57:579–608. - PubMed
-
- Kunkel TA, Erie DA. DNA mismatch repair. Annu Rev Biochem. 2005;74:681–710. - PubMed
-
- Cox EC. Bacterial mutator genes and the control of spontaneous mutation. Annu Rev Genet. 1976;10:135–156. - PubMed
-
- Lynch HT, Smyrk TC, Watson P, Lanspa SJ, Lynch JF, Lynch PM, Cavalieri RJ, Boland CR. Genetics, natural history, tumor spectrum, and pathology of hereditary nonpolyposis colorectal cancer: an updated review. Gastroenterology. 1993;104:1535–1549. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
