

Crossing the species barrier by PrP(Sc) replication in vitro generates unique infectious prions
- PMID: 18775309
- PMCID: PMC2740631
- DOI: 10.1016/j.cell.2008.07.030
Crossing the species barrier by PrP(Sc) replication in vitro generates unique infectious prions
Abstract
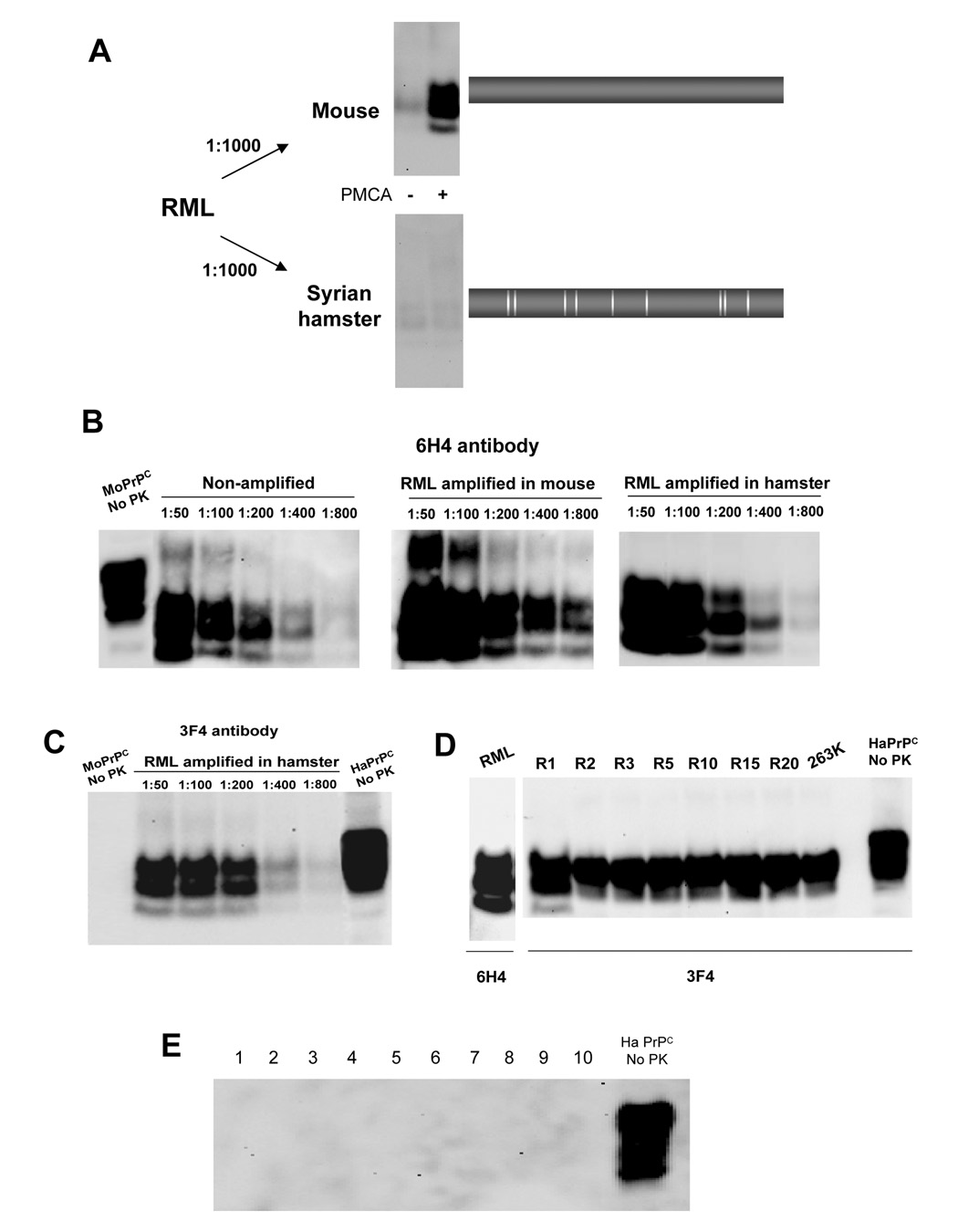
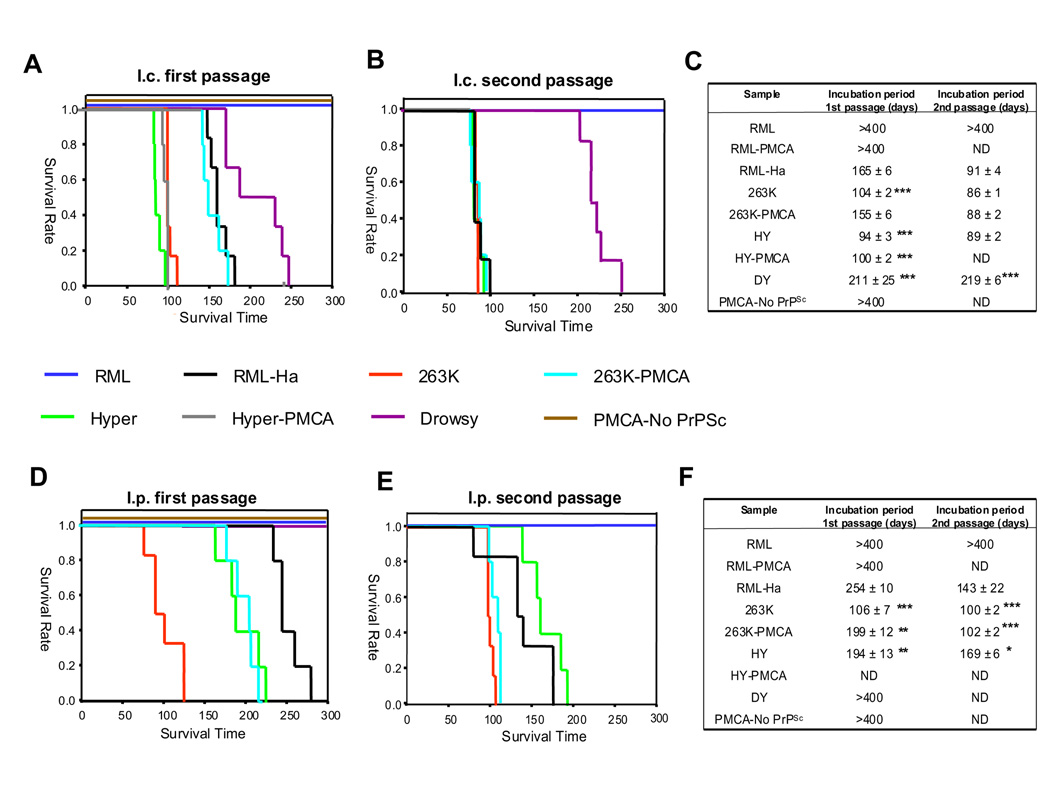
Prions are unconventional infectious agents composed exclusively of misfolded prion protein (PrP(Sc)), which transmits the disease by propagating its abnormal conformation to the cellular prion protein (PrP(C)). A key characteristic of prions is their species barrier, by which prions from one species can only infect a limited number of other species. Here, we report the generation of infectious prions by interspecies transmission of PrP(Sc) misfolding by in vitro PMCA amplification. Hamster PrP(C) misfolded by mixing with mouse PrP(Sc) generated unique prions that were infectious to wild-type hamsters, and similar results were obtained in the opposite direction. Successive rounds of PMCA amplification result in adaptation of the in vitro-produced prions, in a process reminiscent of strain stabilization observed upon serial passage in vivo. Our results indicate that PMCA is a valuable tool for the investigation of cross-species transmission and suggest that species barrier and strain generation are determined by the propagation of PrP misfolding.
Figures


References
-
- Atarashi R, Moore RA, Sim VL, Hughson AG, Dorward DW, Onwubiko HA, Priola SA, Caughey B. Ultrasensitive detection of scrapie prion protein using seeded conversion of recombinant prion protein. Nat. Methods. 2007;4:645–650. - PubMed
-
- Bruce ME. 'New variant' Creutzfeldt-Jakob disease and bovine spongiform encephalopathy. Nat. Med. 2000;6:258–259. - PubMed
-
- Bruce ME. TSE strain variation. Br. Med. Bull. 2003;66:99–108. - PubMed
-
- Buschmann A, Groschup MH. TSE eradication in small ruminants--quo vadis? Berl Munch. Tierarztl. Wochenschr. 2005;118:365–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
