

Prelude to corneal tissue engineering - gaining control of collagen organization
- PMID: 18775789
- PMCID: PMC3712123
- DOI: 10.1016/j.preteyeres.2008.08.001
Prelude to corneal tissue engineering - gaining control of collagen organization
Abstract
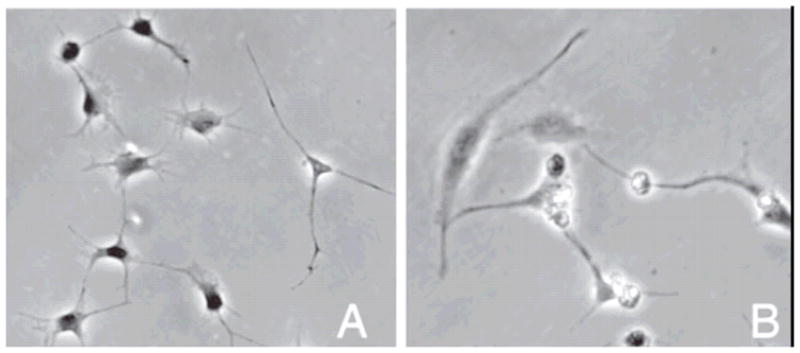
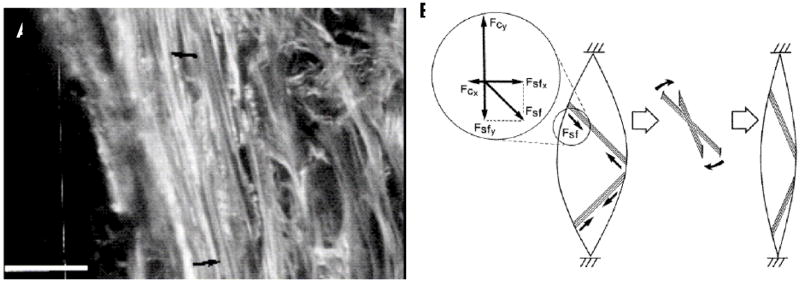
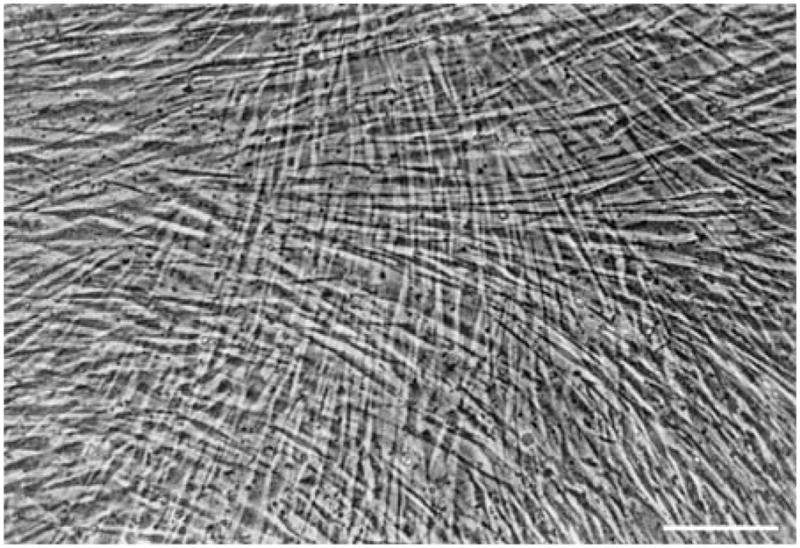
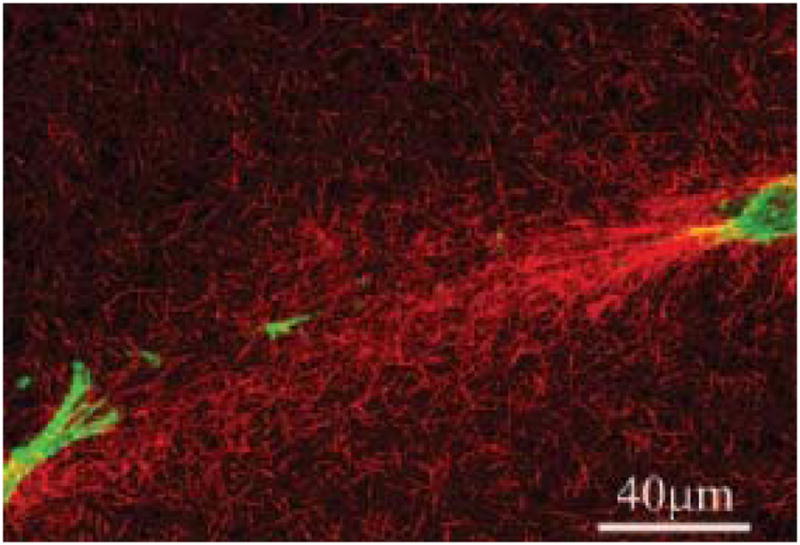
By most standard engineering practice principles, it is premature to credibly discuss the "engineering" of a human cornea. A professional design engineer would assert that we still do not know what a cornea is (and correctly so), therefore we cannot possibly build one. The proof resides in the fact that there are no clinically viable corneas based on classical tissue engineering methods available. This is possibly because tissue engineering in the classical sense (seeding a degradable scaffolding with a population synthetically active cells) does not produce conditions which support the generation of organized tissue. Alternative approaches to the problem are in their infancy and include the methods which attempt to recapitulate development or to produce corneal stromal analogs de novo which require minimal remodeling. Nonetheless, tissue engineering efforts, which have been focused on producing the fundamental functional component of a cornea (organized alternating arrays of collagen or "lamellae"), may have already provided valuable new insights and tools relevant to development, growth, remodeling and pathologies associated with connective tissue in general. This is because engineers ask a fundamentally different question (How can that be done?) than do biological scientists (How is that done?). The difference in inquiry has prompted us to closely examine (and to mimic) development as well as investigate collagen physicochemical behavior so that we may exert control over organization both in cell culture (in vitro) and on the benchtop (de novo). Our initial results indicate that reproducing corneal stroma-like local and long-range organization of collagen may be simpler than we anticipated while controlling spacing and fibril morphology remains difficult, but perhaps not impossible in the (reasonably) near term.
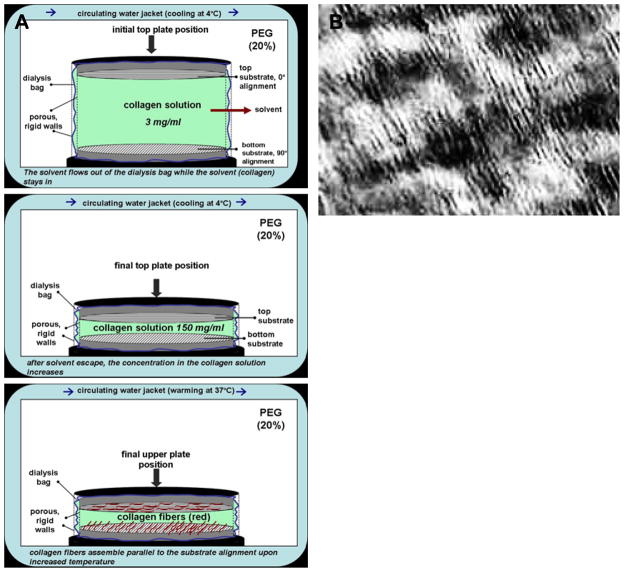
Figures
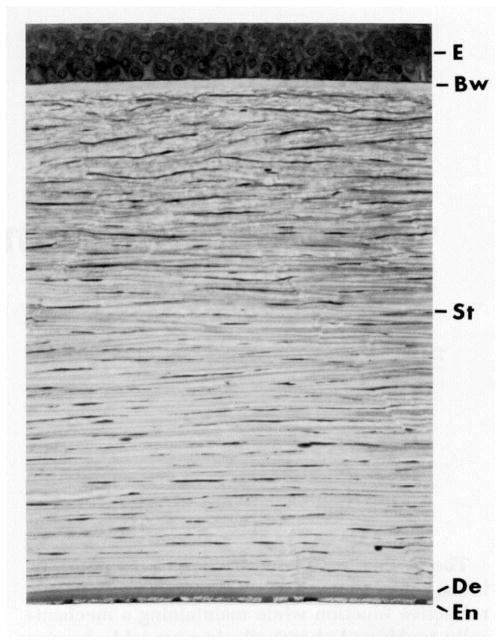
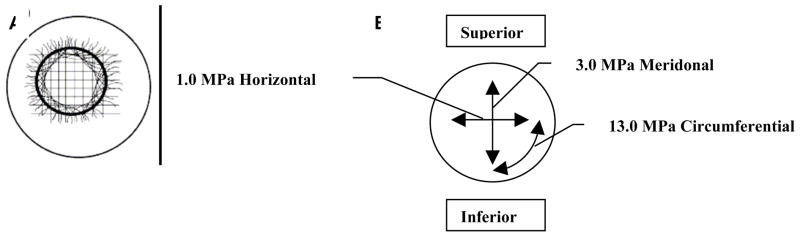


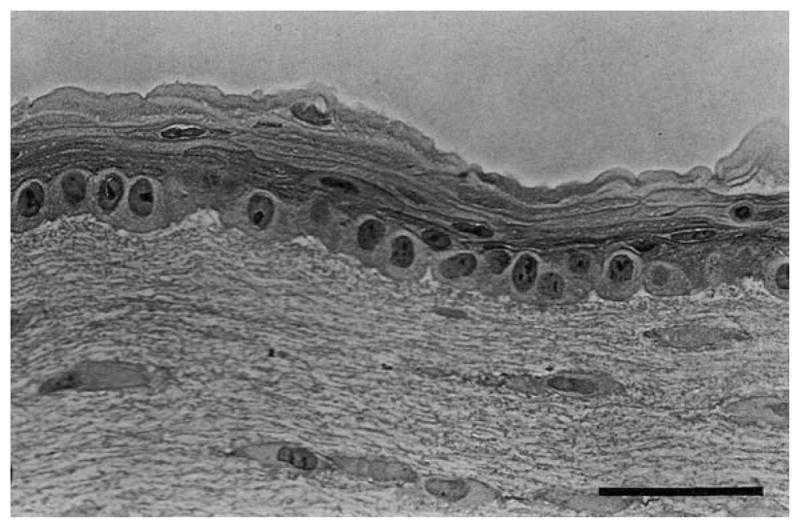
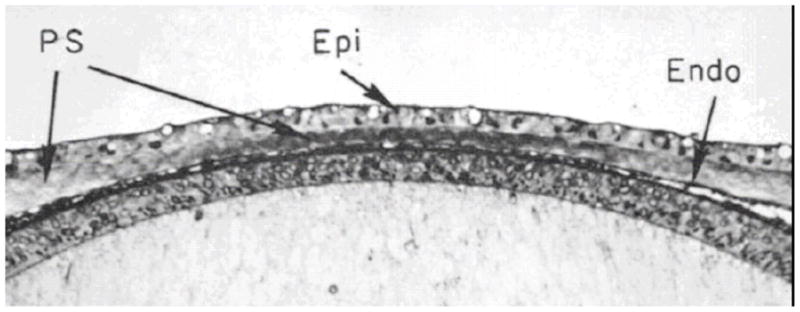

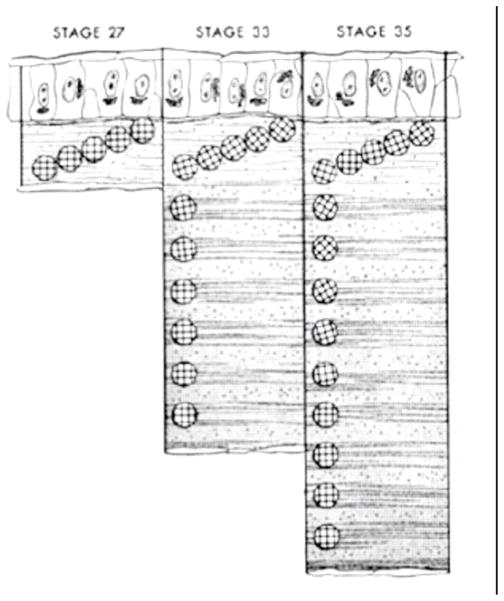
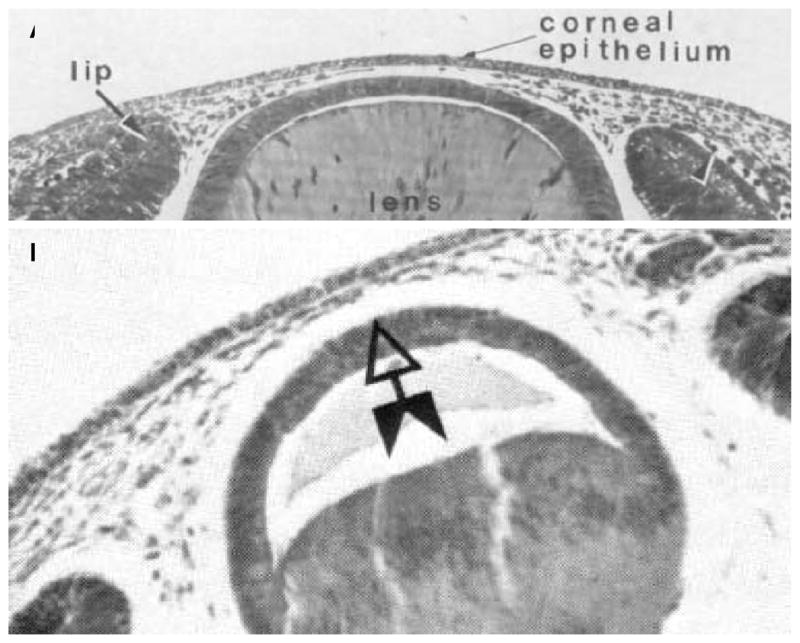

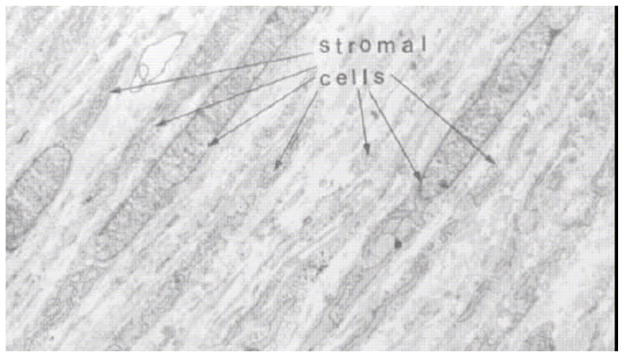
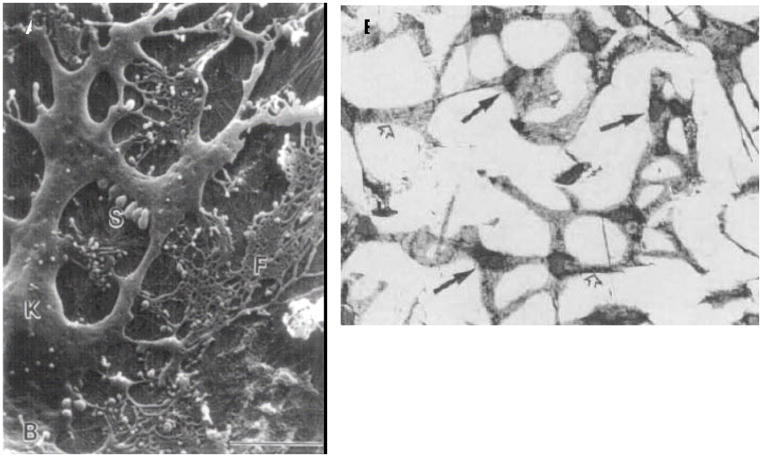

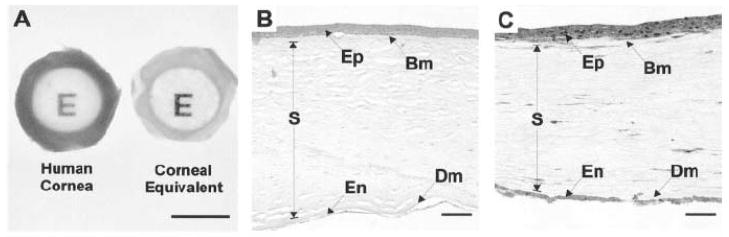
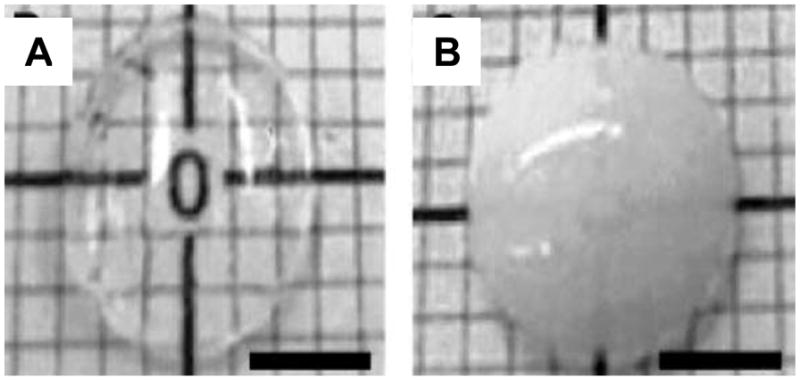
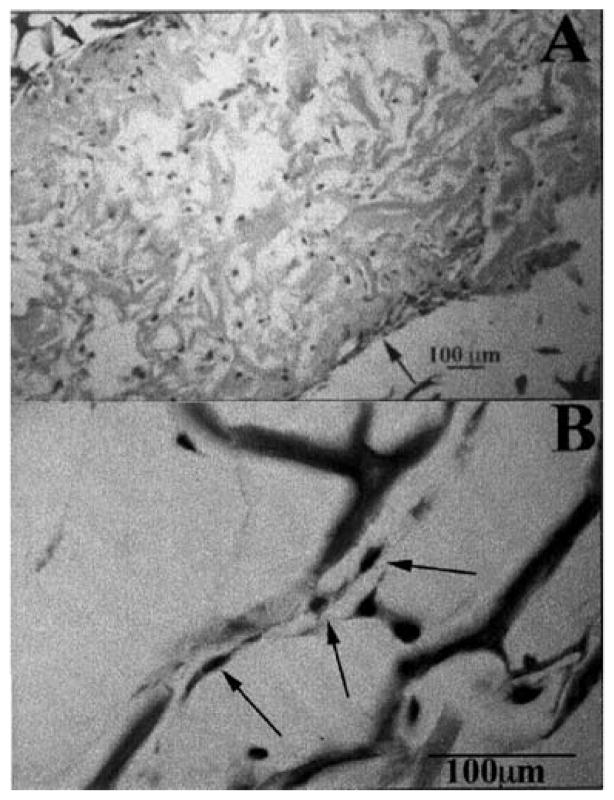
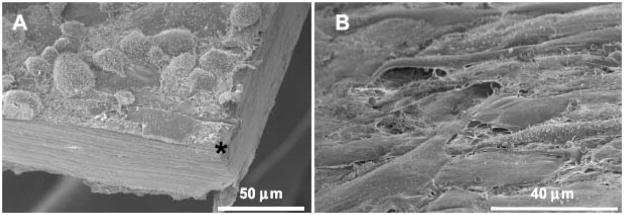
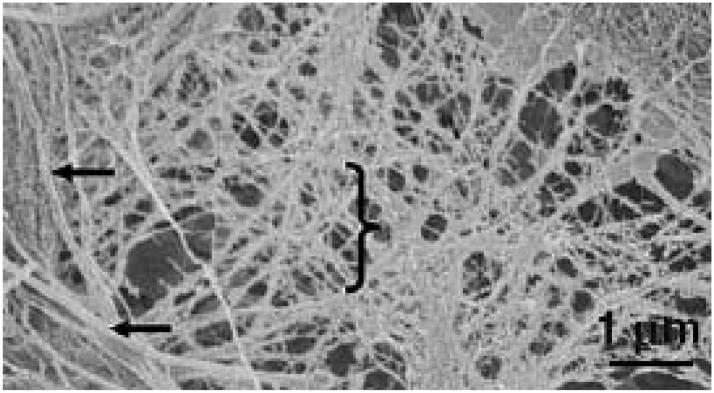




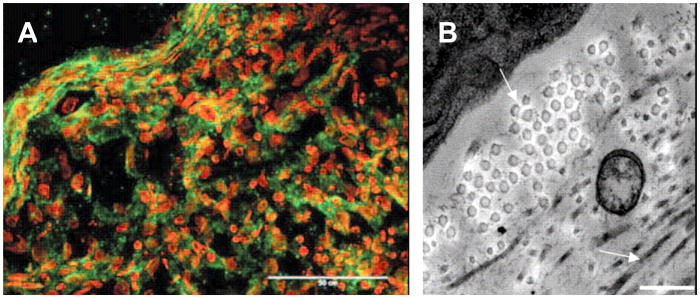
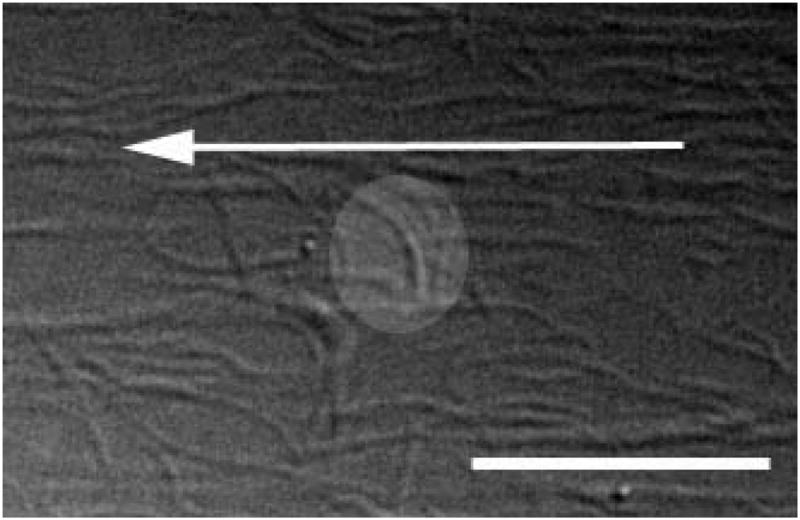
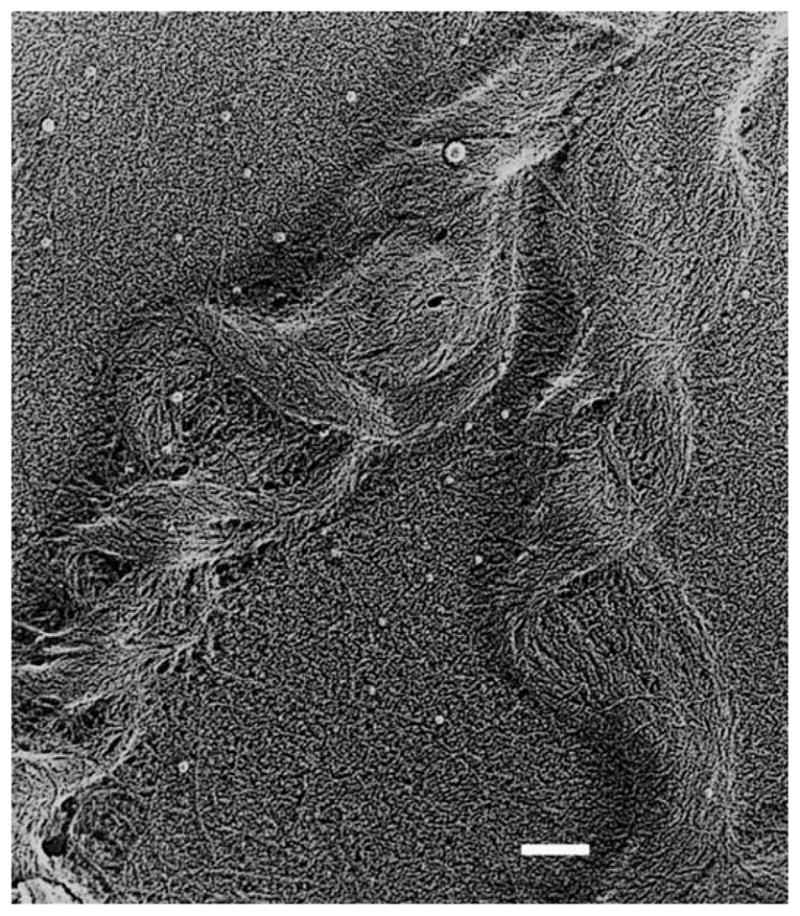
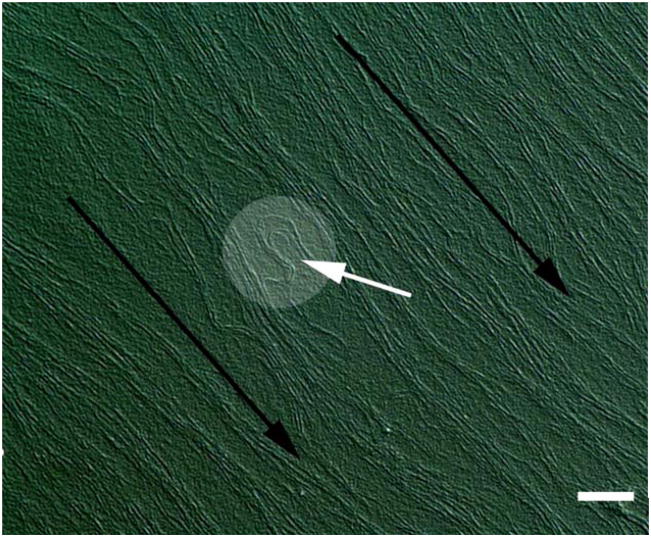
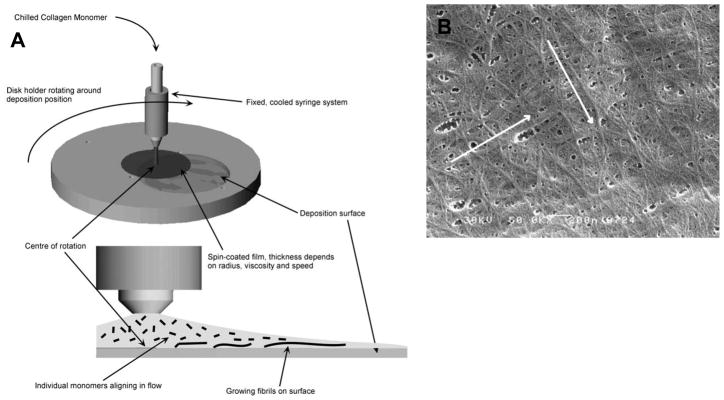
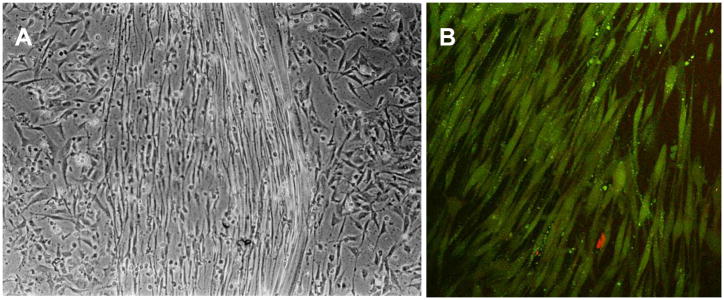
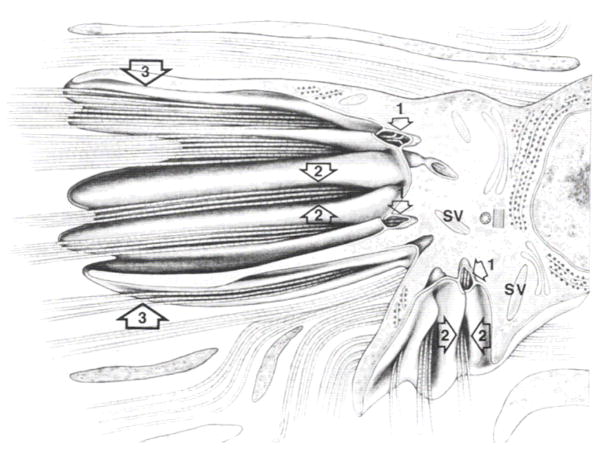
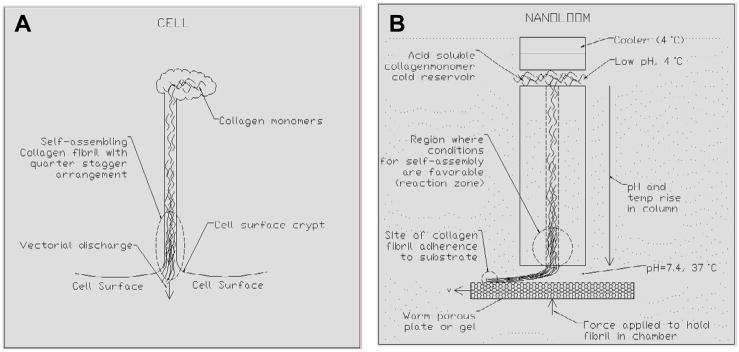
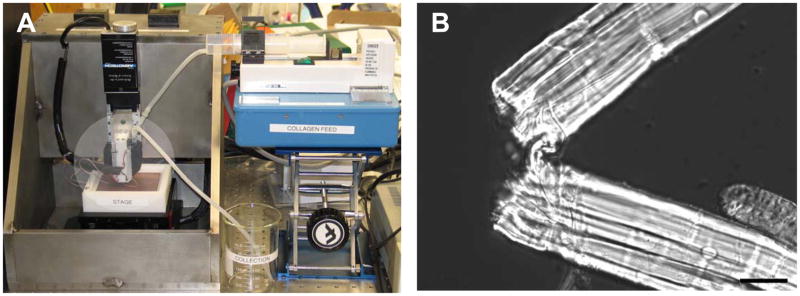
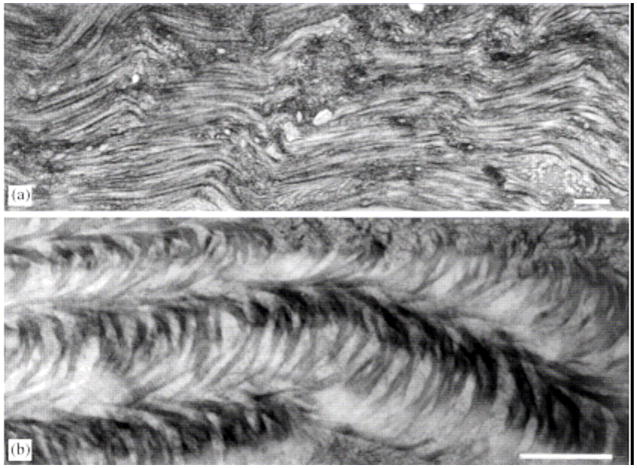
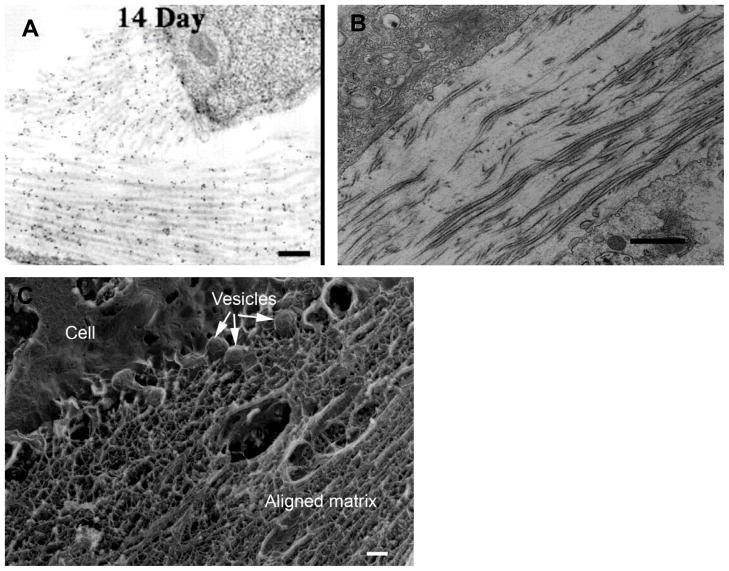


References
-
- Aghamohammadzadeh H, Newton RH, Meek KM. X-ray scattering used to map the preferred collagen orientation in the human cornea and limbus. Structure. 2004;12:249–256. - PubMed
-
- Aiken-O’Neill P, Mannis MJ. Summary of comeal transplant activity Eye Bank Association of America. Cornea. 2002;21:1–3. - PubMed
-
- Akeson WH, Amiel D, Abel MF, Garfin SR, Woo SL. Effects of immobilization on joints. Clin Orthop. 1987:28–37. - PubMed
-
- Altman GH, Horan RL, Martin I, Farhadi J, Stark PR, Volloch V, Richmond JC, Vunjak-Novakovic G, Kaplan DL. Cell differentiation by mechanical stress. Faseb J. 2002;16:270–272. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
