

No requirement for localized Nudel protein expression in Drosophila embryonic axis determination
- PMID: 18776742
- PMCID: PMC2941771
- DOI: 10.4161/fly.6794
No requirement for localized Nudel protein expression in Drosophila embryonic axis determination
Abstract
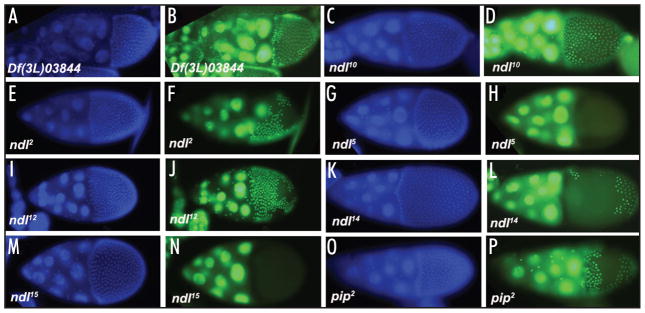
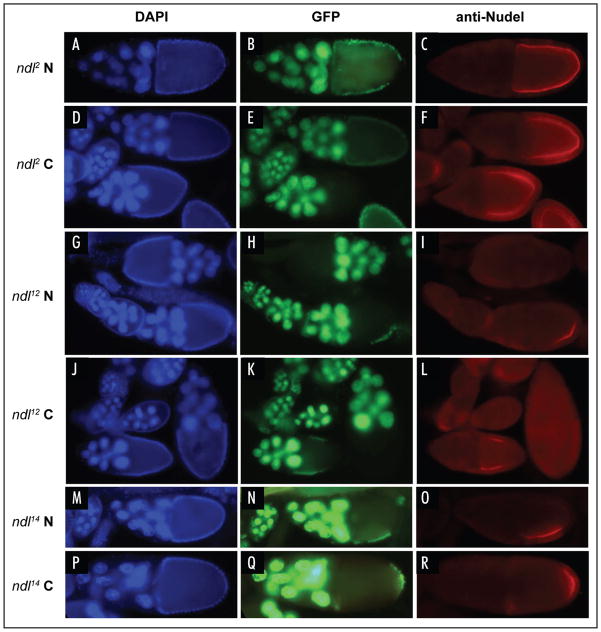
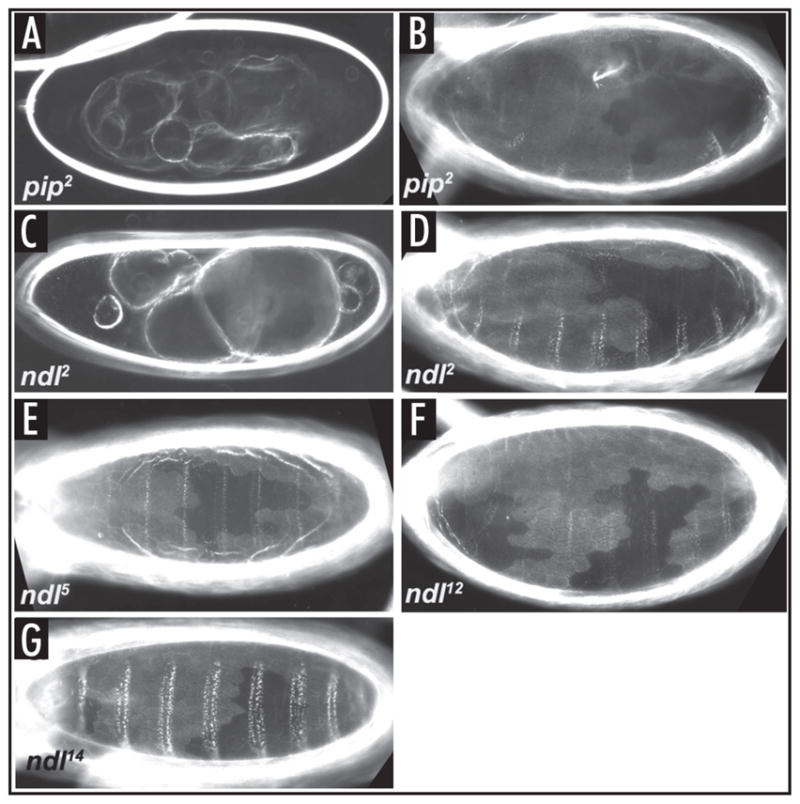
Drosophila embryonic dorsal-ventral polarity is defined by a maternally encoded signal transduction pathway. Gastrulation Defective, Snake, and Easter comprise a serine protease cascade that operates in the perivitelline space to generate active ligand for the Toll receptor, which resides in the embryonic membrane. Toll is activated only on the ventral side of the embryo. Spatial regulation of this pathway is initiated by the ventrally restricted expression of the sulfotransferase Pipe in the follicular epithelium that surrounds the developing oocyte. Pipe is thought to modify a target molecule that is secreted and localized within the ventral region of the egg and future embryo, where it influences the activity of the pathway such that active Toll ligand is produced only ventrally. A potential substrate for Pipe is encoded by nudel, which is expressed throughout the follicle cell layer and encodes a large, multi-functional secreted protein that contains a serine protease domain as well as other structural features characteristic of extracellular matrix proteins. A previous mosaic analysis suggested that the protease domain of Nudel is not a target for Pipe activity as its expression is not required in pipe-expressing cells, but failed to rule out such a role for other functional domains of the protein. To investigate this possibility, we carried out a mosaic analysis of additional nudel alleles, including some that affect the entire protein. Our analysis demonstrated that proteolytically processed segments of Nudel are secreted into the perivitelline space and stably localized, as would be expected for the target of Pipe, However, we found no requirement for nudel to be expressed in ventral, pipe-expressing follicle cells, thereby eliminating Nudel as an essential substrate of Pipe sulfotransferase activity.
Figures
Similar articles
-
Maternal control of the Drosophila dorsal-ventral body axis.Wiley Interdiscip Rev Dev Biol. 2014 Sep-Oct;3(5):301-30. doi: 10.1002/wdev.138. Epub 2014 May 29. Wiley Interdiscip Rev Dev Biol. 2014. PMID: 25124754 Free PMC article. Review.
-
Localized serine protease activity and the establishment of Drosophila embryonic dorsoventral polarity.Fly (Austin). 2013 Jul-Sep;7(3):161-7. doi: 10.4161/fly.25141. Epub 2013 Jun 10. Fly (Austin). 2013. PMID: 24047959 Free PMC article.
-
Activation of Snake in a serine protease cascade that defines the dorsoventral axis is atypical and pipe-independent in Drosophila embryos.FEBS Lett. 2010 Aug 20;584(16):3557-60. doi: 10.1016/j.febslet.2010.07.020. Epub 2010 Jul 16. FEBS Lett. 2010. PMID: 20638387 Free PMC article.
-
A ventrally localized protease in the Drosophila egg controls embryo dorsoventral polarity.Curr Biol. 2012 Jun 5;22(11):1013-8. doi: 10.1016/j.cub.2012.03.065. Epub 2012 May 10. Curr Biol. 2012. PMID: 22578419 Free PMC article.
-
Signaling between somatic follicle cells and the germline patterns the egg and embryo of Drosophila.Curr Top Dev Biol. 2020;140:55-86. doi: 10.1016/bs.ctdb.2019.10.004. Epub 2019 Nov 19. Curr Top Dev Biol. 2020. PMID: 32591083 Review.
Cited by
-
Maternal control of the Drosophila dorsal-ventral body axis.Wiley Interdiscip Rev Dev Biol. 2014 Sep-Oct;3(5):301-30. doi: 10.1002/wdev.138. Epub 2014 May 29. Wiley Interdiscip Rev Dev Biol. 2014. PMID: 25124754 Free PMC article. Review.
References
-
- Moussian B, Roth S. Dorsoventral axis formation in the Drosophila embryo—shaping and transducing a morphogen gradient. Curr Biol. 2005;15:887–99. - PubMed
-
- DeLotto R, Spierer PA. Gene required for the specification of dorsal-ventral pattern in Drosophila appears to encode a serine protease. Nature. 1986;323:688–92. - PubMed
-
- Chasan R, Anderson KV. The role of easter, an apparent serine protease, in organizing the dorsal-ventral pattern of the Drosophila embryo. Cell. 1989;56:391–400. - PubMed
-
- Hong CC, Hashimoto C. An unusual mosaic protein with a protease domain, encoded by the nudel gene, is involved in defining embryonic dorsoventral polarity in Drosophila. Cell. 1995;82:785–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases