

Prevention of methylmercury-induced mitochondrial depolarization, glutathione depletion and cell death by 15-deoxy-delta-12,14-prostaglandin J(2)
- PMID: 18778734
- PMCID: PMC2683755
- DOI: 10.1016/j.neuro.2008.08.003
Prevention of methylmercury-induced mitochondrial depolarization, glutathione depletion and cell death by 15-deoxy-delta-12,14-prostaglandin J(2)
Abstract
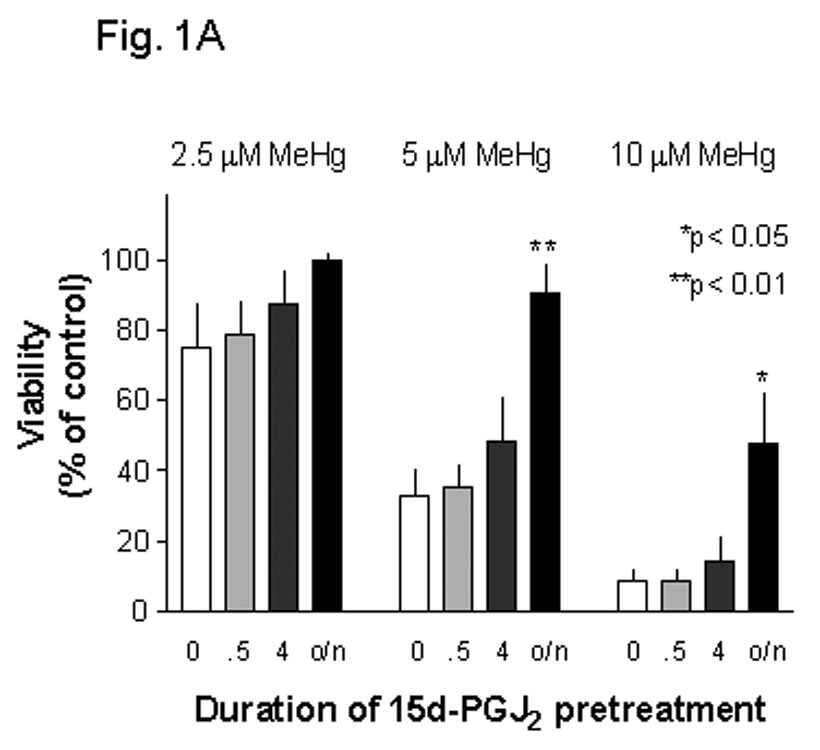
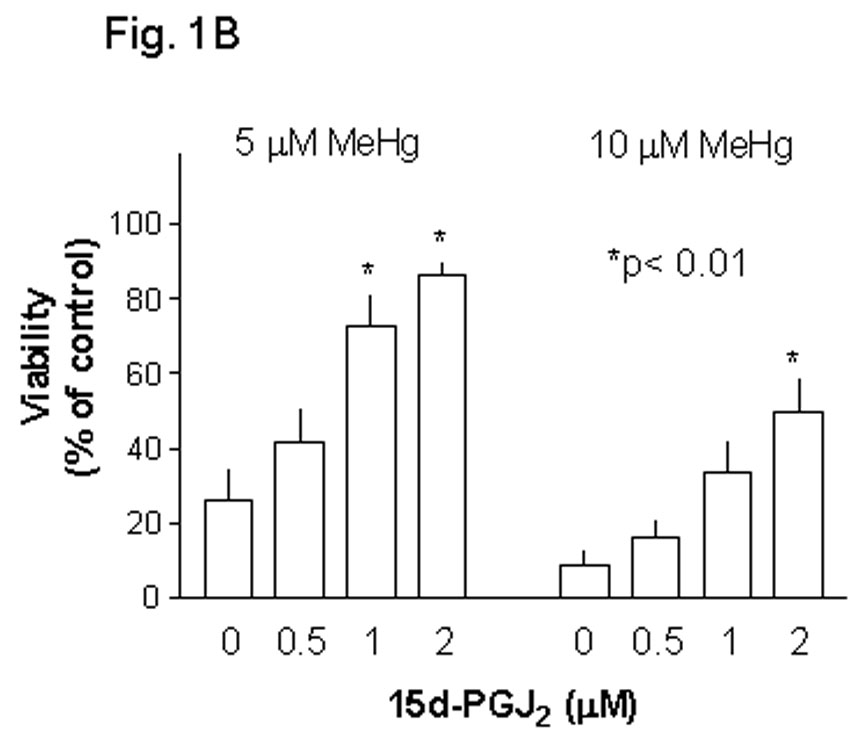
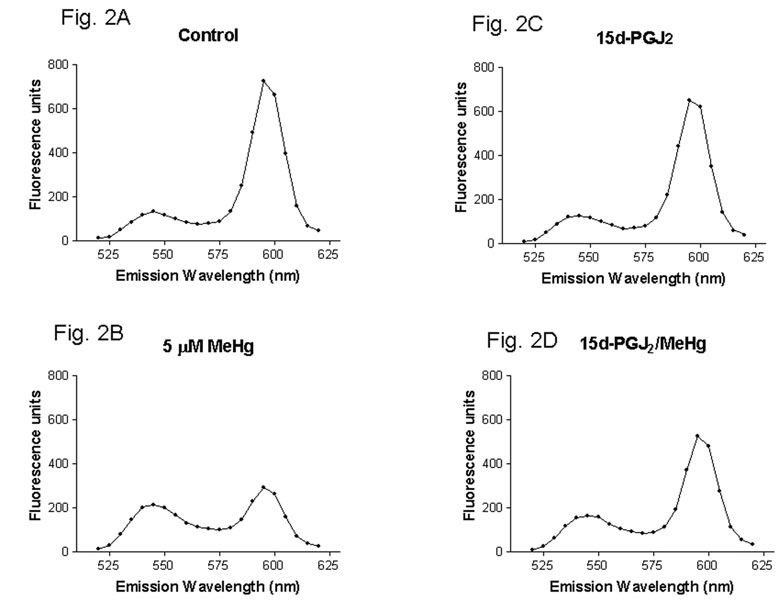
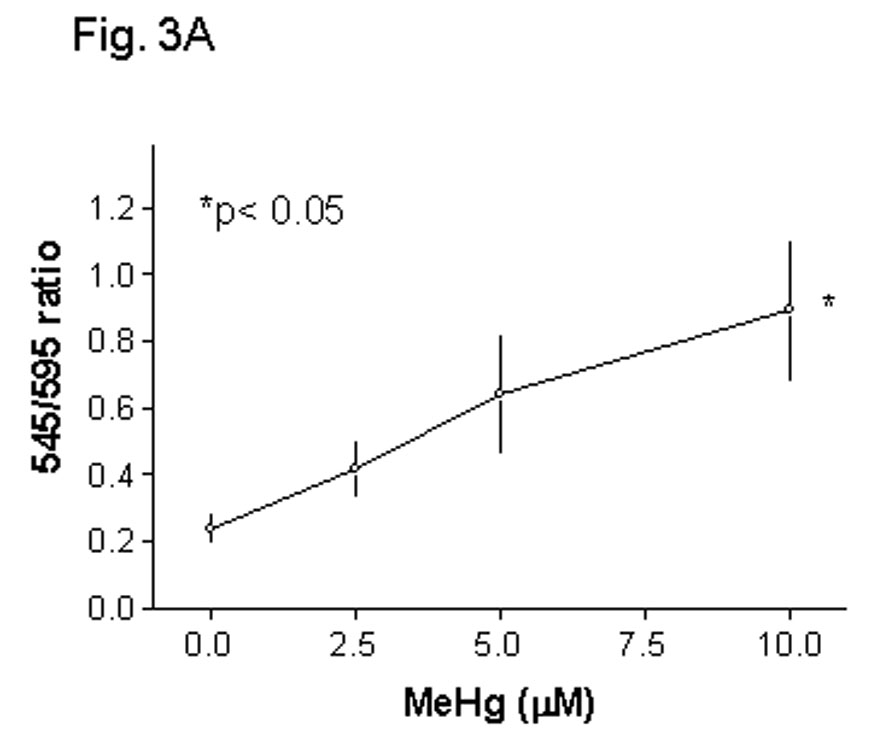
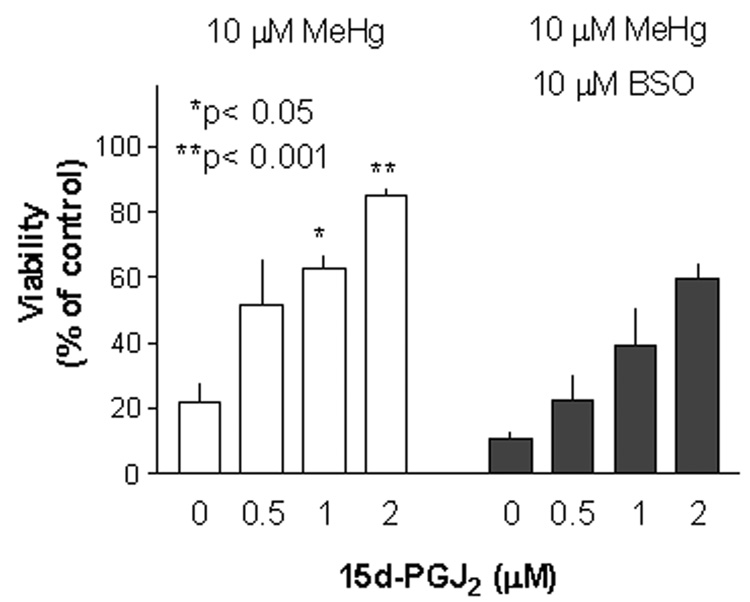
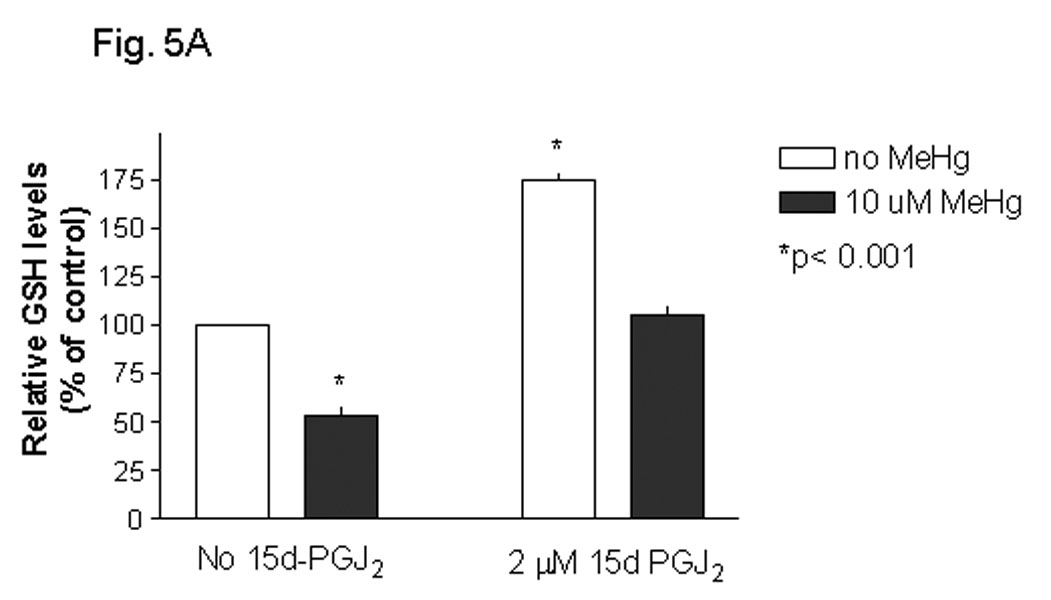
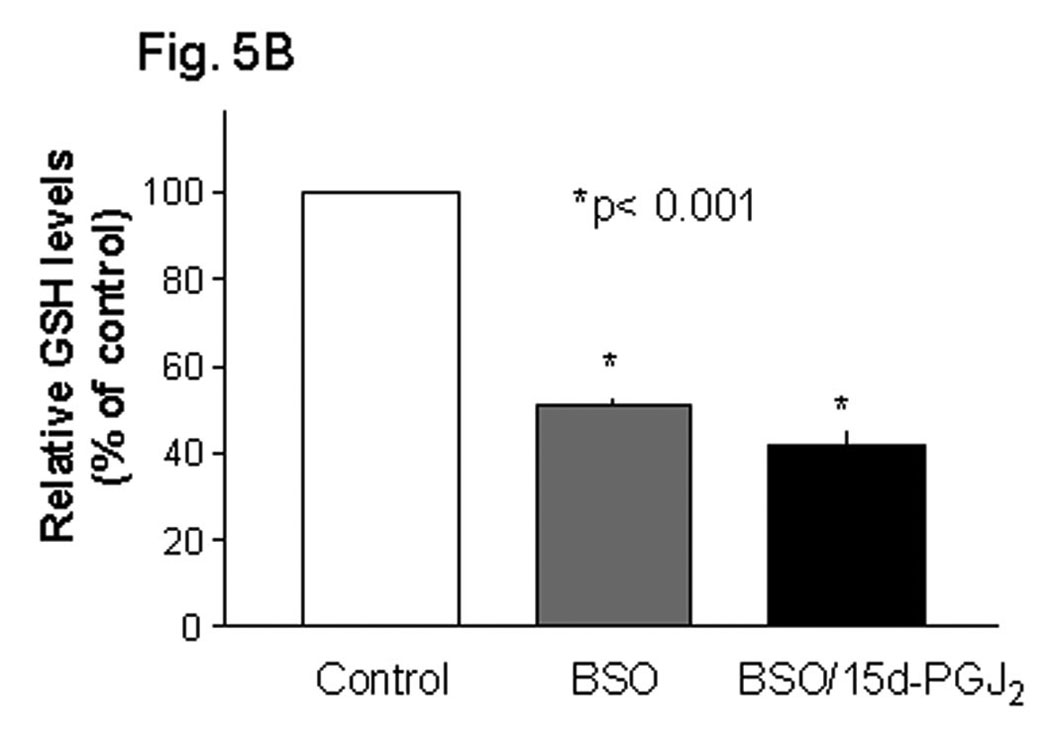
Methylmercury (MeHg) is an environmental toxin that causes severe neurological complications in humans and experimental animals. In addition to neurons, glia in the central nervous system are very susceptible to MeHg toxicity. Pretreatment of glia with the prostaglandin derivative, 15-deoxy-delta-12,14-prostaglandin J(2) (15d-PGJ(2)), caused a significant protection against MeHg cytotoxicity. Results with the C6 glioma cells demonstrated that the protection was dependent on the duration of pretreatment, suggesting that time was required for the up-regulation of cellular defenses. Subsequent experiments indicated that 15d-PGJ(2) prevented MeHg induced mitochondrial depolarization. Similar protection against MeHg cytotoxicity was observed in primary cultures of mouse glia. Analysis of cellular glutathione (GSH) levels indicated that 15d-PGJ(2) caused an up-regulation of GSH and prevented MeHg-induced GSH depletion. Buthionine sulfoximine (BSO), a GSH synthesis inhibitor, completely inhibited the GSH induction by 15d-PGJ(2). However, BSO did not prevent the stabilization of mitochondrial potential and only partially prevented the protection caused by 15d-PGJ(2). While induction of heme oxygenase-1 was implicated in the cytoprotection by 15d-PGJ(2) under some experimental conditions, additional experiments indicated that this enzyme was not involved in the cytoprotection observed in this system. Together, these results suggested that while up-regulation of GSH by 15d-PGJ(2) might help cells to defend against MeHg toxicity, there may be other yet unidentified mechanism(s) initiated by 15d-PGJ(2) treatment that contributed to its protection against MeHg cytotoxicity.
Figures


Similar articles
-
Protective effects of 15-deoxy-Delta12,14-prostaglandin J2 against glutamate-induced cell death in primary cortical neuron cultures: induction of adaptive response and enhancement of cell tolerance primarily through up-regulation of cellular glutathione.J Neurochem. 2007 Sep;102(5):1625-1634. doi: 10.1111/j.1471-4159.2007.04701.x. J Neurochem. 2007. PMID: 17697048
-
15-Deoxy-delta12,14-prostaglandin-J(2) up-regulates cyclooxygenase-2 but inhibits prostaglandin-E(2) production through a thiol antioxidant-sensitive mechanism.Pharmacol Res. 2008 May;57(5):344-50. doi: 10.1016/j.phrs.2008.03.007. Epub 2008 Mar 26. Pharmacol Res. 2008. PMID: 18456507
-
Modulatory effect of glutathione status and antioxidants on methylmercury-induced free radical formation in primary cultures of cerebral astrocytes.Brain Res Mol Brain Res. 2005 Jun 13;137(1-2):11-22. doi: 10.1016/j.molbrainres.2005.02.006. Epub 2005 Mar 17. Brain Res Mol Brain Res. 2005. PMID: 15950756
-
Biphasic effects of 15-deoxy-delta(12,14)-prostaglandin J(2) on glutathione induction and apoptosis in human endothelial cells.Arterioscler Thromb Vasc Biol. 2001 Nov;21(11):1846-51. doi: 10.1161/hq1101.098488. Arterioscler Thromb Vasc Biol. 2001. PMID: 11701476
-
The role of intracellular glutathione in methylmercury-induced toxicity in embryonic neuronal cells.Neurotoxicology. 1999 Oct;20(5):793-804. Neurotoxicology. 1999. PMID: 10591515
Cited by
-
Morphine protects against methylmercury intoxication: a role for opioid receptors in oxidative stress?PLoS One. 2014 Oct 17;9(10):e110815. doi: 10.1371/journal.pone.0110815. eCollection 2014. PLoS One. 2014. PMID: 25330195 Free PMC article.
-
Maternal thimerosal exposure results in aberrant cerebellar oxidative stress, thyroid hormone metabolism, and motor behavior in rat pups; sex- and strain-dependent effects.Cerebellum. 2012 Jun;11(2):575-86. doi: 10.1007/s12311-011-0319-5. Cerebellum. 2012. PMID: 22015705
-
Modulation of methylmercury uptake by methionine: prevention of mitochondrial dysfunction in rat liver slices by a mimicry mechanism.Toxicol Appl Pharmacol. 2011 Apr 1;252(1):28-35. doi: 10.1016/j.taap.2011.01.010. Epub 2011 Jan 27. Toxicol Appl Pharmacol. 2011. PMID: 21276810 Free PMC article.
-
Antioxidant effects of Dendropanax morbifera Léveille extract in the hippocampus of mercury-exposed rats.BMC Complement Altern Med. 2015 Jul 23;15:247. doi: 10.1186/s12906-015-0786-1. BMC Complement Altern Med. 2015. PMID: 26201852 Free PMC article.
-
Sulforaphane Prevents Methylmercury-Induced Oxidative Damage and Excitotoxicity Through Activation of the Nrf2-ARE Pathway.Mol Neurobiol. 2017 Jan;54(1):375-391. doi: 10.1007/s12035-015-9643-y. Epub 2016 Jan 7. Mol Neurobiol. 2017. PMID: 26742517
References
-
- Allen JW, El-Oqayli H, Aschner M, Syversen T, Sonnewald U. Methylmercury has a selective effect on mitochondria in cultured astrocytes in the presence of [U-(13)C]glutamate. Brain Res. 2001;908:149–154. - PubMed
-
- Alvarez-Maqueda M, El Bekay R, Alba G, et al. 15-deoxy-delta 12,14-prostaglandin J2 induces heme oxygenase-1 gene expression in a reactive oxygen species-dependent manner in human lymphocytes. J Biol Chem. 2004;279:21929–21937. - PubMed
-
- Aoun P, Simpkins JW, Agarwal N. Role of PPAR-gamma ligands in neuroprotection against glutamate-induced cytotoxicity in retinal ganglion cells. Invest Ophthalmol Vis Sci. 2003;44:2999–3004. - PubMed
-
- Aschner M. Methylmercury in astrocytes--what possible significance? Neurotoxicology. 1996;17:93–106. - PubMed
-
- Aschner M, Allen JW, Kimelberg HK, LoPachin RM, Streit WJ. Glial cells in neurotoxicity development. Annu Rev Pharmacol Toxicol. 1999;39:151–173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
