

Acyclovir is activated into a HIV-1 reverse transcriptase inhibitor in herpesvirus-infected human tissues
- PMID: 18779052
- PMCID: PMC4210193
- DOI: 10.1016/j.chom.2008.07.008
Acyclovir is activated into a HIV-1 reverse transcriptase inhibitor in herpesvirus-infected human tissues
Abstract
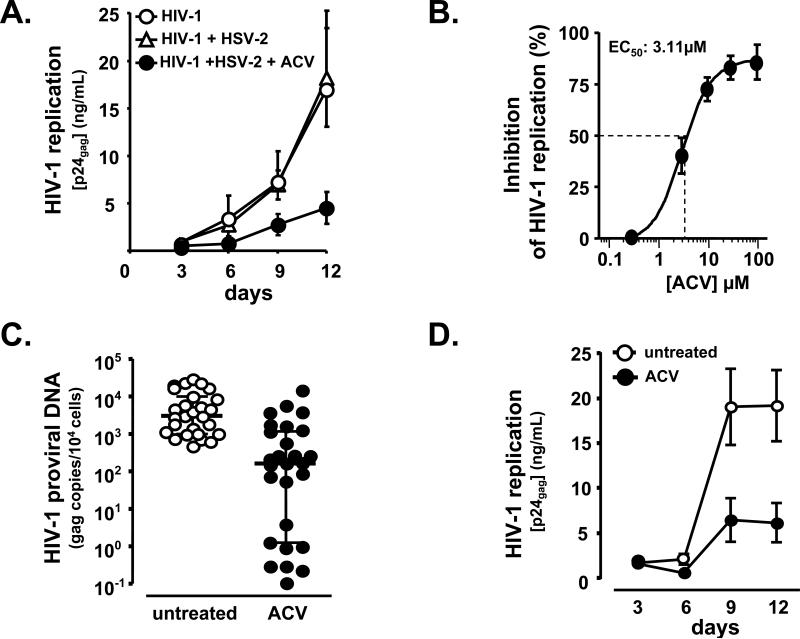
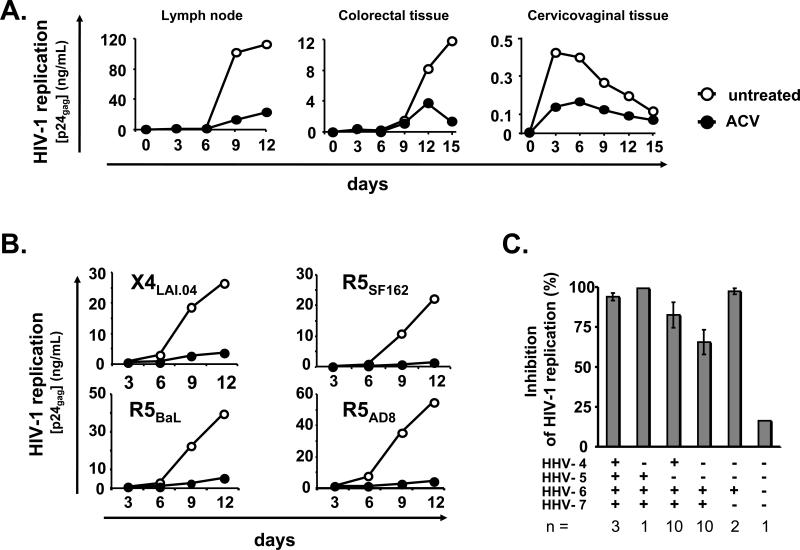
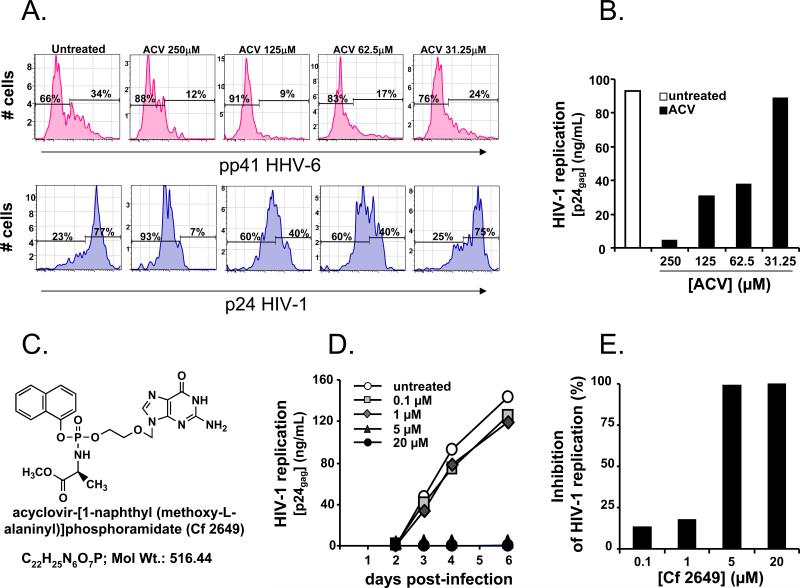
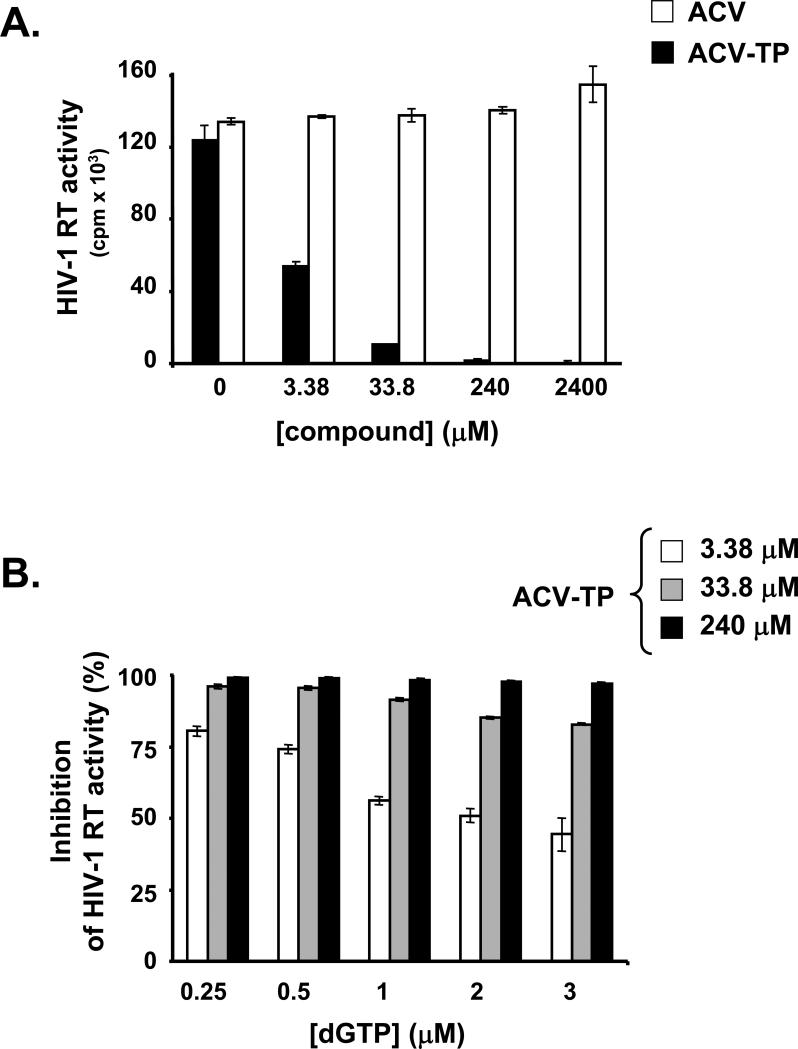
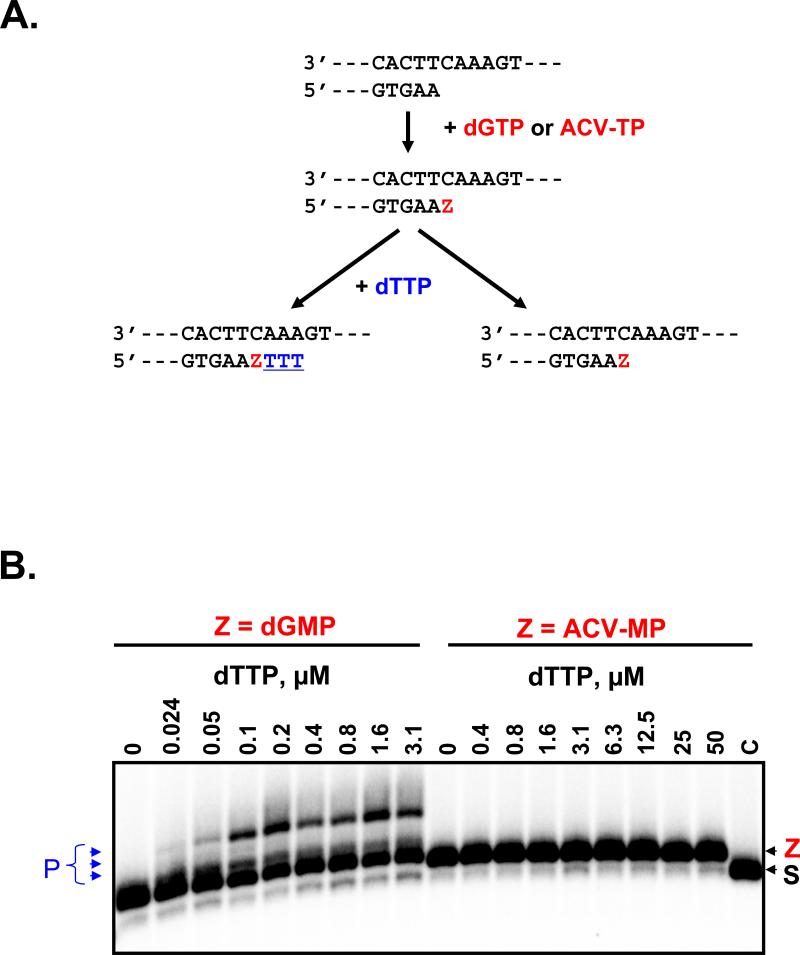
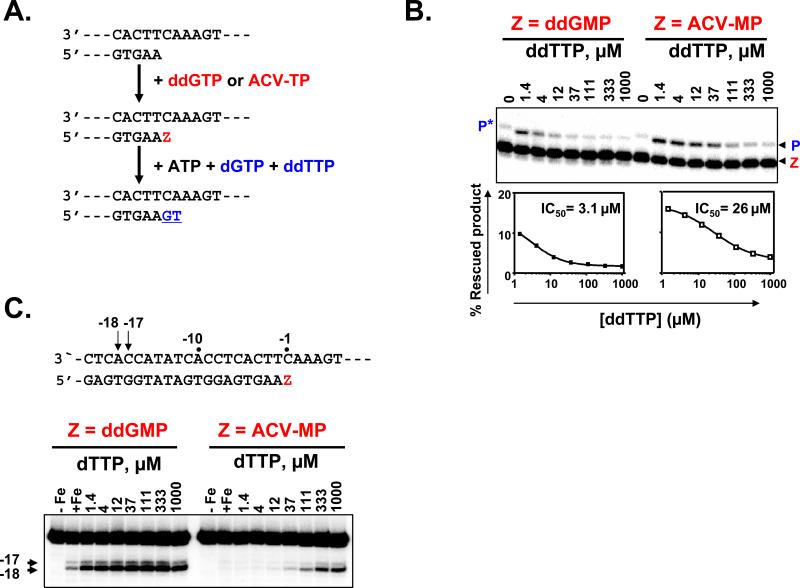
For most viruses, there is a need for antimicrobials that target unique viral molecular properties. Acyclovir (ACV) is one such drug. It is activated into a human herpesvirus (HHV) DNA polymerase inhibitor exclusively by HHV kinases and, thus, does not suppress other viruses. Here, we show that ACV suppresses HIV-1 in HHV-coinfected human tissues, but not in HHV-free tissue or cell cultures. However, addition of HHV-6-infected cells renders these cultures sensitive to anti-HIV ACV activity. We hypothesized that such HIV suppression requires ACV phosphorylation by HHV kinases. Indeed, an ACV monophosphorylated prodrug bypasses the HHV requirement for HIV suppression. Furthermore, phosphorylated ACV directly inhibits HIV-1 reverse transcriptase (RT), terminating DNA chain elongation, and can trap RT at the termination site. These data suggest that ACV anti-HIV-1 activity may contribute to the response of HIV/HHV-coinfected patients to ACV treatment and could guide strategies for the development of new HIV-1 RT inhibitors.
Figures
Comment in
-
Coinfection alters the playing field: herpesviruses induce acyclovir to inhibit HIV.Cell Host Microbe. 2008 Sep 11;4(3):194-5. doi: 10.1016/j.chom.2008.08.006. Cell Host Microbe. 2008. PMID: 18779044
References
-
- Delta Coordinating Committee and Delta Virology Committee HIV-1 RNA response to antiretroviral treatment in 1280 participants in the Delta Trial: an extended virology study. Aids. 1999;13:57–65. - PubMed
-
- Baeten JM, S. L., Lucchetti A, Whittington WLH, Sanchez J, Coombs RW, Margaret A, Wald A, Corey L, Celum C. Herpes simplex virus suppressive treatment decreases plasma HIV-1 load in HSV-2/HIV-1 co-infected: a randomised, placebo-controlled, cross-over trial. 17th International Society fpr Sexually Transmitted Diseases Research Meeting; Seattle, WA. 2007.
-
- Barral K, Hider RC, Balzarini J, Neyts J, De Clercq E, Camplo M. Synthesis and antiviral evaluation of 3-hydroxy-2-methylpyridin-4-one dideoxynucleoside derivatives. Bioorg Med Chem Lett. 2003;13:4371–4374. - PubMed
-
- Burrows FJ, Gore M, Smiley WR, Kanemitsu MY, Jolly DJ, Read SB, Nicholas T, Kruse CA. Purified herpes simplex virus thymidine kinase retroviral particles: III. Characterization of bystander killing mechanisms in transfected tumor cells. Cancer Gene Ther. 2002;9:87–95. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical