

Role of a pro-sequence in the secretory pathway of prothyrotropin-releasing hormone
- PMID: 18779326
- PMCID: PMC2581549
- DOI: 10.1074/jbc.M803413200
Role of a pro-sequence in the secretory pathway of prothyrotropin-releasing hormone
Abstract
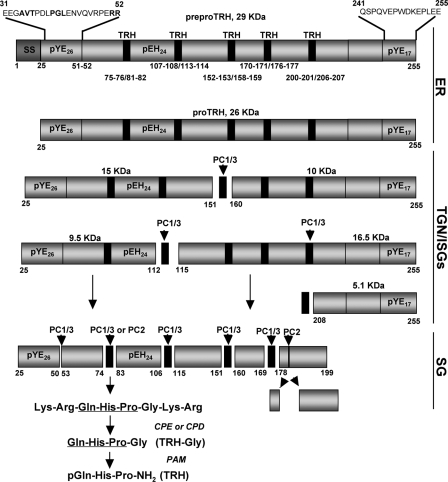
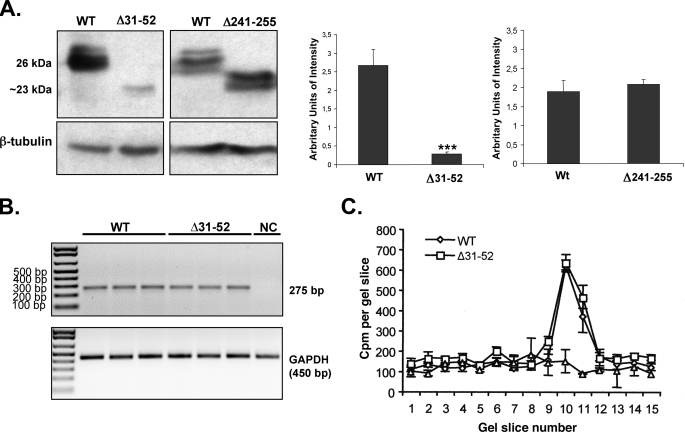
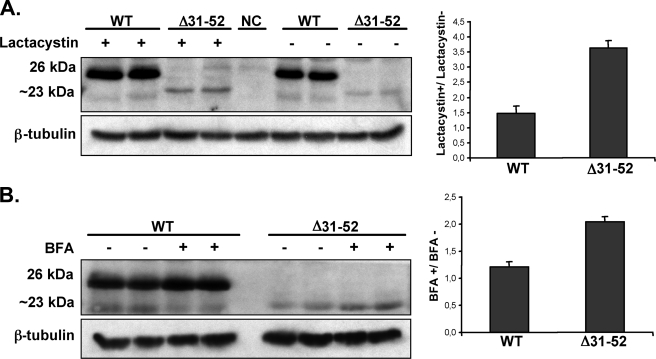
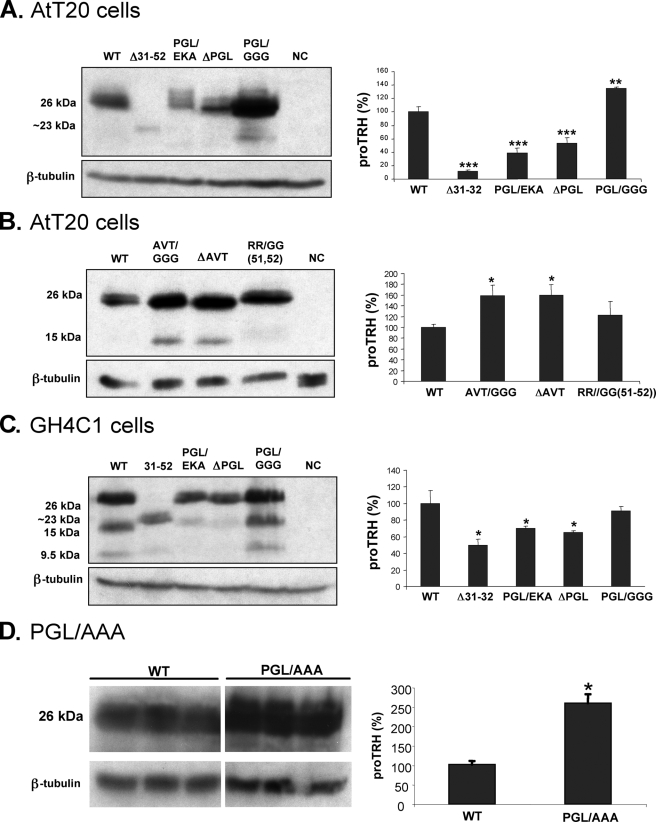
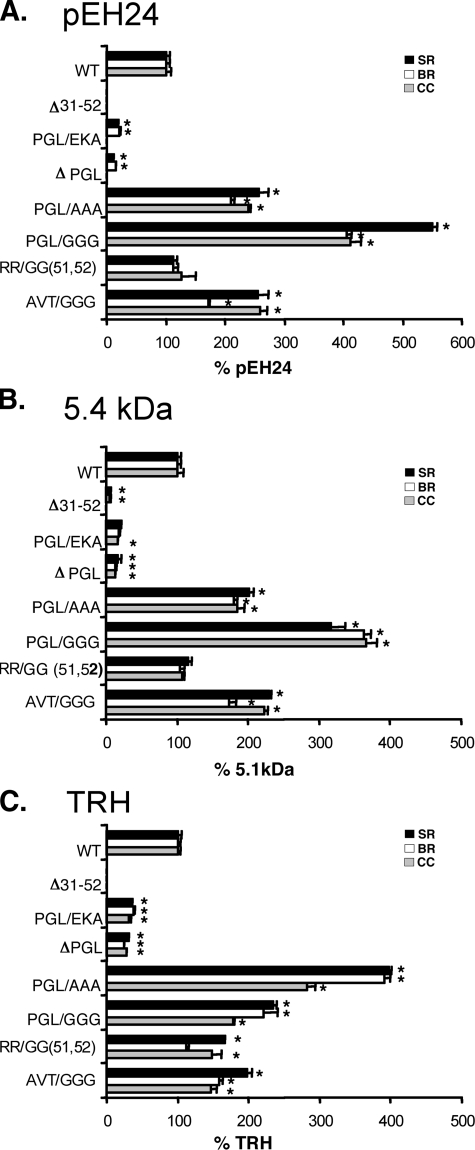
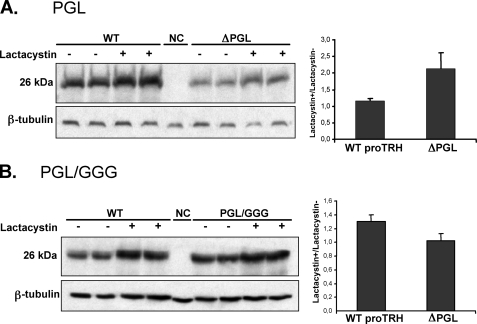
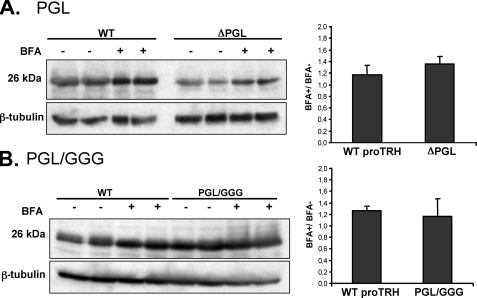
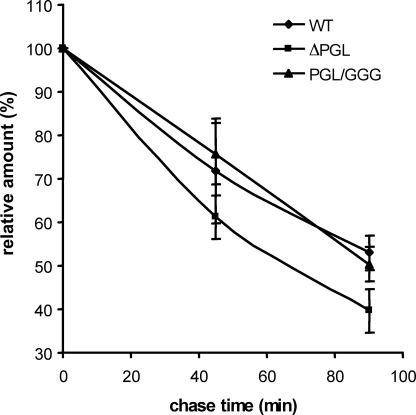
The biogenesis of rat thyrotropin releasing hormone (TRH) involves the processing of its precursor (proTRH) into five biologically active TRH peptides and several non-TRH peptides where two of them had been attributed potential biological functions. This process implicates 1) proper folding of proTRH in the endoplasmic reticulum after its biosynthesis and exit to the Golgi apparatus and beyond, 2) initial processing of proTRH in the trans Golgi network and, 3) sorting of proTRH-derived peptides to the regulated secretory pathway. Previous studies have focused on elucidating the processing and sorting determinants of proTRH. However, the role of protein folding in the sorting of proTRH remains unexplored. Here we have investigated the role in the secretion of proTRH of a sequence comprising 22 amino acid residues, located at the N-terminal region of proTRH, residues 31-52. Complete deletion of these 22 amino acids dramatically compromised the biosynthesis of proTRH, manifested as a severe reduction in the steady state level of proTRH in the endoplasmic reticulum. This effect was largely reproduced by the deletion of only three amino acid residues, 40PGL42, within the proTRH31-52 sequence. The decreased steady state level of the mutant DeltaPGL was due to enhanced endoplasmic reticulum-associated protein degradation. However, the remnant of DeltaPGL that escaped degradation was properly processed and sorted to secretory granules. Thus, these results suggest that the N-terminal domain within the prohormone sequence does not act as "sorting signal" in late secretion; instead, it seems to play a key role determining the proper folding pathway of the precursor and, thus, its stability.
Figures
References
-
- Nillni, E. A., and Sevarino, K. A (1999) Endocr. Rev. 20 599–648 - PubMed
-
- Zhou, A., Bloomquist, B. T., and Mains, R. E. (1993) J. Biol. Chem. 268 1763–1769 - PubMed
-
- Steiner, D. F. (1998) Curr. Opin. Chem. Biol. 2 31–39 - PubMed
-
- Seidah, N. G., and Chretien, M. (1999) Brain Res. 848 45–62 - PubMed
-
- Scamuffa, N., Calvo, F., Chretien, M., Seidah, N. G., and Khatib, A. M. (2006) FASEB J. 20 1954–1963 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
