

Outer surface protein A protects Lyme disease spirochetes from acquired host immunity in the tick vector
- PMID: 18779341
- PMCID: PMC2573341
- DOI: 10.1128/IAI.00410-08
Outer surface protein A protects Lyme disease spirochetes from acquired host immunity in the tick vector
Abstract
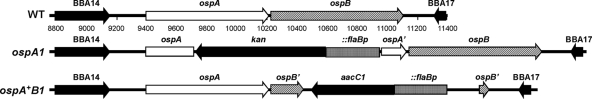
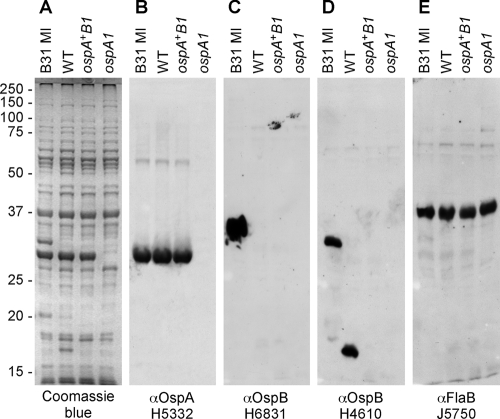
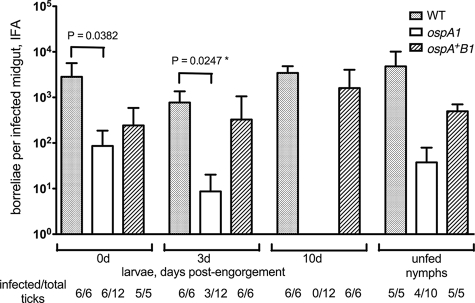
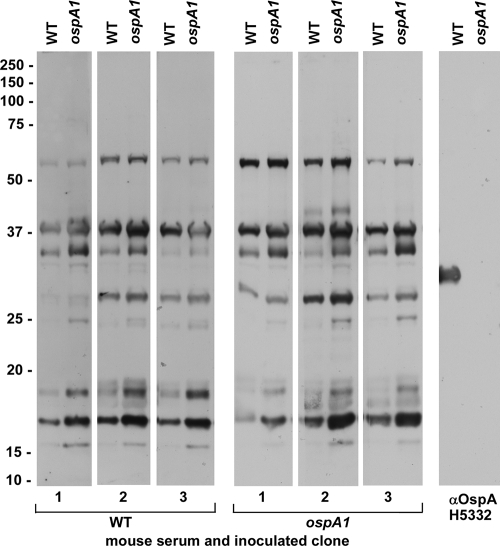
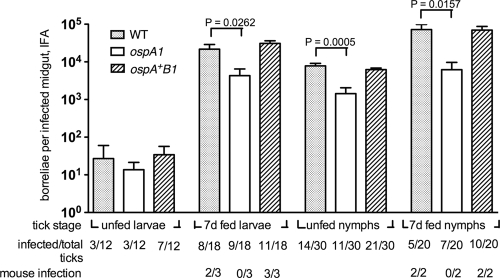
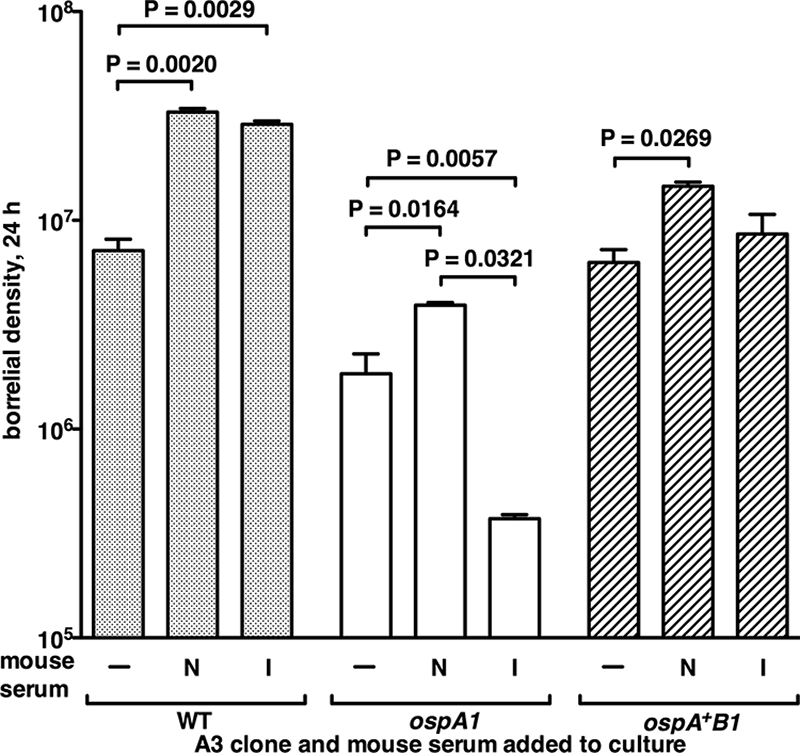
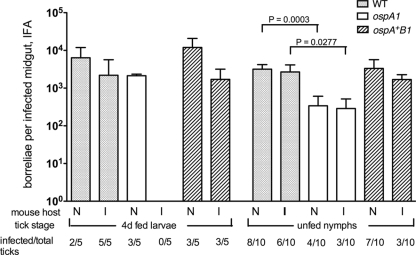
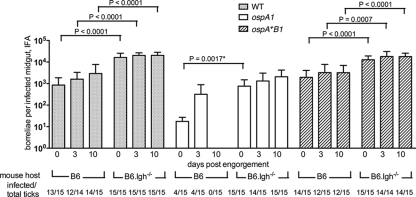
The Lyme disease spirochete Borrelia burgdorferi alters the expression of outer surface protein (osp) genes as the bacterium cycles between ticks and mammals. OspA is produced as borreliae enter the tick vector and remains a major surface antigen during midgut colonization. To elucidate the role of OspA in the vector, we created an insertional deletion of ospA in strain B31-A3. The ospA mutant infects mice when it is injected intradermally and is acquired by larval ticks fed on these mice, where it persists through the molt to the nymph stage. Bacterial survival rates in artificially infected tick larvae fed on naïve mice were compared with those in the vector fed on immune mice. The ospA mutant proliferates in larvae if it is exposed to blood from naïve mice, but it declines in density after larval feeding if the blood is from immune mice. When uninfected larvae are fed on B-cell-deficient mice infected with the ospA mutant, larvae show borrelial densities and persistence that are significantly greater than those fed on infected, immunocompetent mice. We conclude that OspA serves a critical antibody-shielding role during vector blood meal uptake from immune hosts and is not required for persistence in the tick vector.
Figures
References
-
- Barbour, A. G., and C. F. Garon. 1987. Linear plasmids of the bacterium Borrelia burgdorferi have covalently closed ends. Science 237409-411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
