

Physiological significance of a peripheral tissue circadian clock
- PMID: 18779586
- PMCID: PMC2532700
- DOI: 10.1073/pnas.0806717105
Physiological significance of a peripheral tissue circadian clock
Abstract
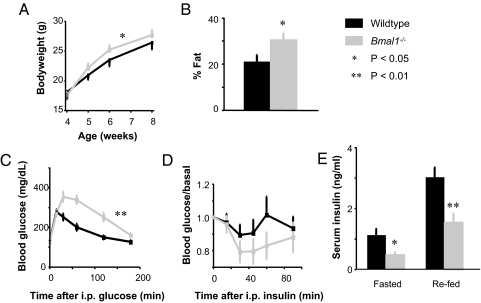
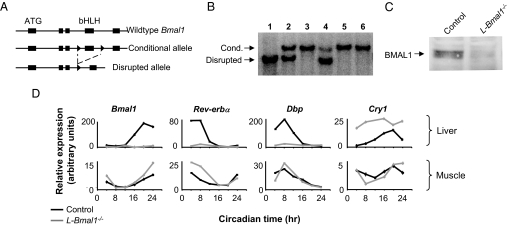
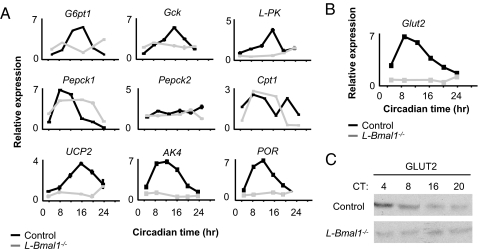
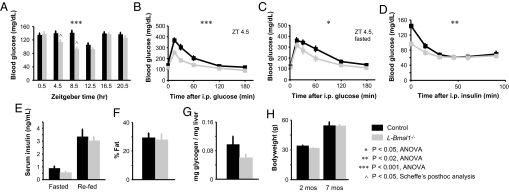
Mammals have circadian clocks in peripheral tissues, but there is no direct evidence of their physiological importance. Unlike the suprachiasmatic nucleus clock that is set by light and drives rest-activity and fasting-feeding cycles, peripheral clocks are set by daily feeding, suggesting that at least some contribute metabolic regulation. The liver plays a well known role in glucose homeostasis, and we report here that mice with a liver-specific deletion of Bmal1, an essential clock component, exhibited hypoglycemia restricted to the fasting phase of the daily feeding cycle, exaggerated glucose clearance, and loss of rhythmic expression of hepatic glucose regulatory genes. We conclude that the liver clock is important for buffering circulating glucose in a time-of-day-dependent manner. Our findings suggest that the liver clock contributes to homeostasis by driving a daily rhythm of hepatic glucose export that counterbalances the daily cycle of glucose ingestion resulting from the fasting-feeding cycle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Circadian glucose homeostasis requires compensatory interference between brain and liver clocks.Proc Natl Acad Sci U S A. 2008 Sep 30;105(39):14753-4. doi: 10.1073/pnas.0807861105. Epub 2008 Sep 23. Proc Natl Acad Sci U S A. 2008. PMID: 18812506 Free PMC article. No abstract available.
References
-
- Young MW, Kay SA. Time zones: A comparative genetics of circadian clocks. Nat Rev Genet. 2001;2:702–715. - PubMed
-
- Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum Mol Genet. 2006;15:R271–R277. - PubMed
-
- Tosini G, Menaker M. Circadian rhythms in cultured mammalian retina. Science. 1996;272:419–421. - PubMed
-
- Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in cultured Rat-1 fibroblasts. Cell. 1998;93:929–937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
