

MAVS and MyD88 are essential for innate immunity but not cytotoxic T lymphocyte response against respiratory syncytial virus
- PMID: 18780793
- PMCID: PMC2532974
- DOI: 10.1073/pnas.0804717105
MAVS and MyD88 are essential for innate immunity but not cytotoxic T lymphocyte response against respiratory syncytial virus
Abstract
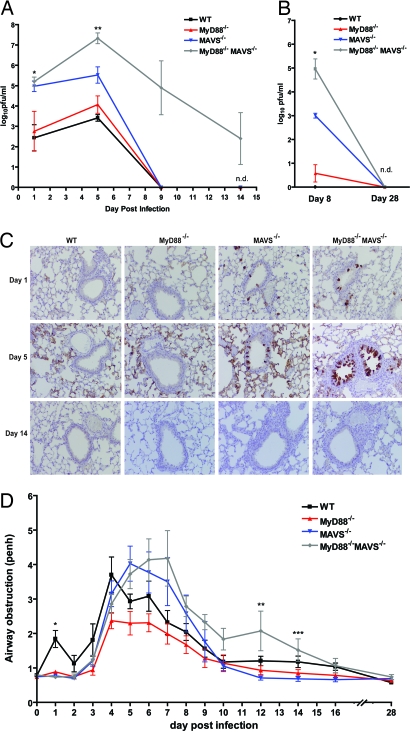
Infection by RNA viruses is detected by the host through Toll-like receptors or RIG-I-like receptors. Toll-like receptors and RIG-I-like receptors signal through the adaptors MyD88 and MAVS, respectively, to induce type I IFNs (IFN-I) and other antiviral molecules, which are thought to be essential for activating the adaptive immune system. We investigated the role of these adaptors in innate and adaptive immune responses against respiratory syncytial virus (RSV), a common human pathogen. Deletion of Mavs abolished the induction of IFN-I and other proinflammatory cytokines by RSV. Genome-wide expression profiling in the lung showed that the vast majority of RSV-induced genes depended on MAVS. Although Myd88 deficiency did not affect most RSV-induced genes, mice lacking both adaptors harbored a higher and more prolonged viral load and exhibited more severe pulmonary disease than those lacking either adaptor alone. Surprisingly, Myd88(-/-)Mavs(-/-) mice were able to activate a subset of pulmonary dendritic cells that traffic to the draining lymph node in response to RSV. These mice subsequently mounted a normal cytotoxic T-lymphocyte response and demonstrated delayed but effective viral clearance. These results provide an example of a normal and effective adaptive immune response in the absence of innate immunity mediated by MAVS and MyD88.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Pichlmair A, Reis e Sousa C. Innate recognition of viruses. Immunity. 2007;27:370–383. - PubMed
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Seth RB, Sun L, Chen ZJ. Antiviral innate immunity pathways. Cell Res. 2006;16:141–147. - PubMed
-
- Pichlmair A, et al. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science. 2006;314:997–1001. - PubMed
-
- Hornung V, et al. 5′-Triphosphate RNA is the ligand for RIG-I. Science. 2006;314:994–997. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
