

The human mitochondrial ribosome recycling factor is essential for cell viability
- PMID: 18782833
- PMCID: PMC2566884
- DOI: 10.1093/nar/gkn576
The human mitochondrial ribosome recycling factor is essential for cell viability
Abstract
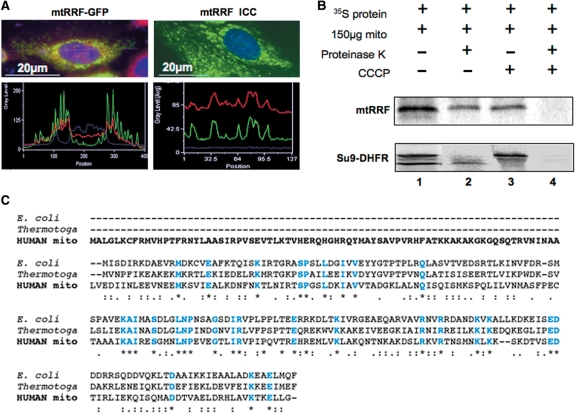
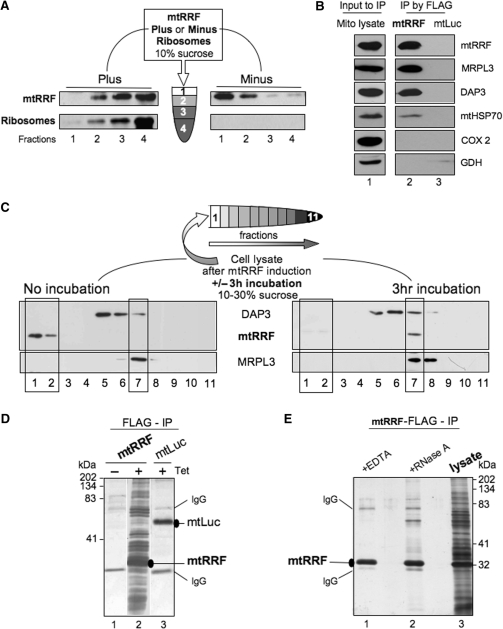
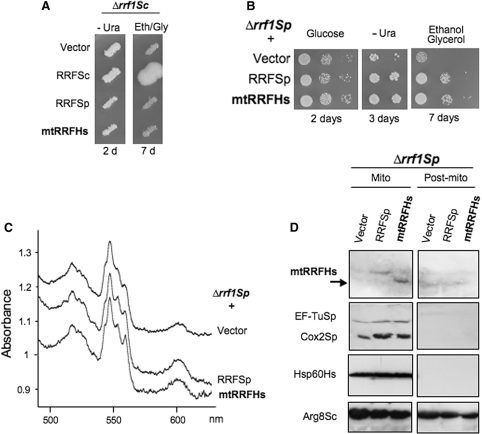
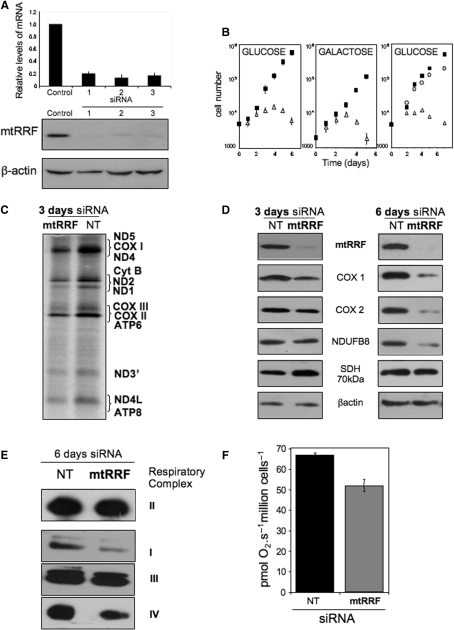
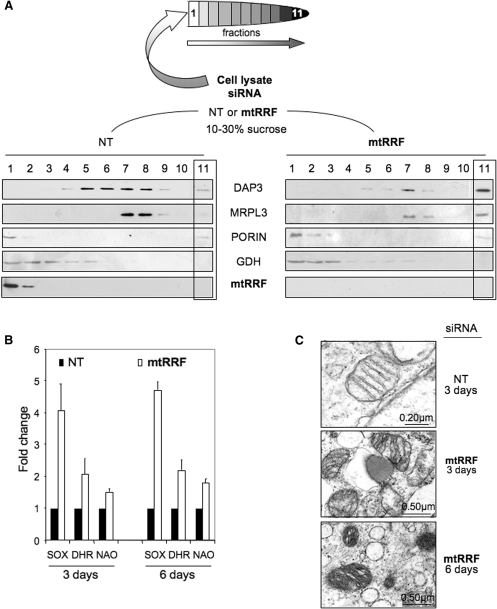
The molecular mechanism of human mitochondrial translation has yet to be fully described. We are particularly interested in understanding the process of translational termination and ribosome recycling in the mitochondrion. Several candidates have been implicated, for which subcellular localization and characterization have not been reported. Here, we show that the putative mitochondrial recycling factor, mtRRF, is indeed a mitochondrial protein. Expression of human mtRRF in fission yeast devoid of endogenous mitochondrial recycling factor suppresses the respiratory phenotype. Further, human mtRRF is able to associate with Escherichia coli ribosomes in vitro and can associate with mitoribosomes in vivo. Depletion of mtRRF in human cell lines is lethal, initially causing profound mitochondrial dysmorphism, aggregation of mitoribosomes, elevated mitochondrial superoxide production and eventual loss of OXPHOS complexes. Finally, mtRRF was shown to co-immunoprecipitate a large number of mitoribosomal proteins attached to other mitochondrial proteins, including putative members of the mitochondrial nucleoid.
Figures
Similar articles
-
Structural basis of translation termination, rescue, and recycling in mammalian mitochondria.Mol Cell. 2021 Jun 17;81(12):2566-2582.e6. doi: 10.1016/j.molcel.2021.03.042. Epub 2021 Apr 19. Mol Cell. 2021. PMID: 33878294
-
The Insertion Domain of Mti2 Facilitates the Association of Mitochondrial Initiation Factors with Mitoribosomes in Schizosaccharomyces pombe.Biomolecules. 2025 May 10;15(5):695. doi: 10.3390/biom15050695. Biomolecules. 2025. PMID: 40427588 Free PMC article.
-
Essential function of Aco2, a fusion protein of aconitase and mitochondrial ribosomal protein bL21, in mitochondrial translation in fission yeast.FEBS Lett. 2015 Mar 24;589(7):822-8. doi: 10.1016/j.febslet.2015.02.015. Epub 2015 Feb 25. FEBS Lett. 2015. PMID: 25724335
-
Maintaining mitochondrial ribosome function: The role of ribosome rescue and recycling factors.RNA Biol. 2022;19(1):117-131. doi: 10.1080/15476286.2021.2015561. Epub 2021 Dec 31. RNA Biol. 2022. PMID: 34923906 Free PMC article. Review.
-
Supernumerary proteins of mitochondrial ribosomes.Biochim Biophys Acta. 2014 Apr;1840(4):1227-32. doi: 10.1016/j.bbagen.2013.08.010. Epub 2013 Aug 17. Biochim Biophys Acta. 2014. PMID: 23958563 Review.
Cited by
-
Biological significance of 5S rRNA import into human mitochondria: role of ribosomal protein MRP-L18.Genes Dev. 2011 Jun 15;25(12):1289-305. doi: 10.1101/gad.624711. Genes Dev. 2011. PMID: 21685364 Free PMC article.
-
Illuminating mitochondrial translation through mouse models.Hum Mol Genet. 2024 May 22;33(R1):R61-R79. doi: 10.1093/hmg/ddae020. Hum Mol Genet. 2024. PMID: 38779771 Free PMC article. Review.
-
Does mtDNA nucleoid organization impact aging?Exp Gerontol. 2010 Aug;45(7-8):473-7. doi: 10.1016/j.exger.2009.12.002. Epub 2009 Dec 11. Exp Gerontol. 2010. PMID: 20004238 Free PMC article. Review.
-
Polymorphisms in the mitochondrial ribosome recycling factor EF-G2mt/MEF2 compromise cell respiratory function and increase atorvastatin toxicity.PLoS Genet. 2012;8(6):e1002755. doi: 10.1371/journal.pgen.1002755. Epub 2012 Jun 14. PLoS Genet. 2012. PMID: 22719265 Free PMC article.
-
Mitochondrial OXPHOS Biogenesis: Co-Regulation of Protein Synthesis, Import, and Assembly Pathways.Int J Mol Sci. 2020 May 28;21(11):3820. doi: 10.3390/ijms21113820. Int J Mol Sci. 2020. PMID: 32481479 Free PMC article. Review.
References
-
- Anderson S, Bankier AT, Barrell BG, De Bruijn MHL, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, et al. Sequence and organization of the human mitochondrial genome. Nature. 1981;290:457–465. - PubMed
-
- Pel HJ, Grivell LA. Protein synthesis in mitochondria. Mol. Biol. Rep. 1994;19:183–194. - PubMed
-
- Chung HK, Spremulli LL. Purification and characterization of elongation factor G from bovine liver mitochondria. J. Biol. Chem. 1990;265:21000–21004. - PubMed
-
- Koc EC, Spremulli LL. Identification of mammalian mitochondrial translational initiation factor 3 and examination of its role in initiation complex formation with natural mRNAs. J. Biol. Chem. 2002;277:35541–35549. - PubMed
-
- Schwartzbach CJ, Spremulli LL. Bovine mitochondrial protein synthesis elongation factors. Identification and initial characterization of an elongation factor Tu-elongation factor Ts complex. J. Biol. Chem. 1989;264:19125–19131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
