

Crystal structure of human plasma platelet-activating factor acetylhydrolase: structural implication to lipoprotein binding and catalysis
- PMID: 18784071
- PMCID: PMC2581546
- DOI: 10.1074/jbc.M804750200
Crystal structure of human plasma platelet-activating factor acetylhydrolase: structural implication to lipoprotein binding and catalysis
Abstract
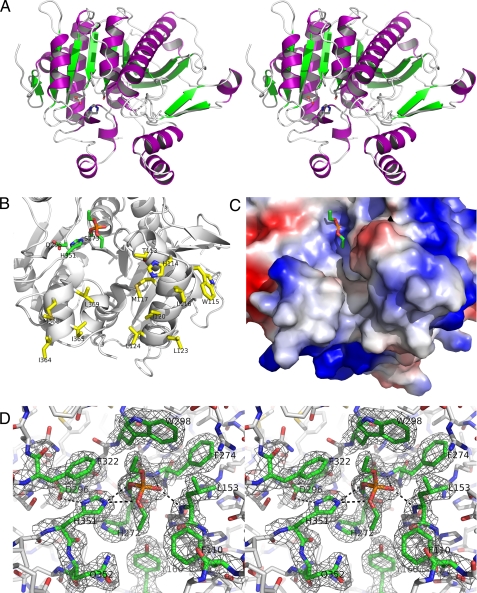
Human plasma platelet-activating factor (PAF) acetylhydrolase functions by reducing PAF levels as a general anti-inflammatory scavenger and is linked to anaphylactic shock, asthma, and allergic reactions. The enzyme has also been implicated in hydrolytic activities of other pro-inflammatory agents, such as sn-2 oxidatively fragmented phospholipids. This plasma enzyme is tightly bound to low and high density lipoprotein particles and is also referred to as lipoprotein-associated phospholipase A2. The crystal structure of this enzyme has been solved from x-ray diffraction data collected to a resolution of 1.5 angstroms. It has a classic lipase alpha/beta-hydrolase fold, and it contains a catalytic triad of Ser273, His351, and Asp296. Two clusters of hydrophobic residues define the probable interface-binding region, and a prediction is given of how the enzyme is bound to lipoproteins. Additionally, an acidic patch of 10 carboxylate residues and a neighboring basic patch of three residues are suggested to play a role in high density lipoprotein/low density lipoprotein partitioning. A crystal structure is also presented of PAF acetylhydrolase reacted with the organophosphate compound paraoxon via its active site Ser273. The resulting diethyl phosphoryl complex was used to model the tetrahedral intermediate of the substrate PAF to the active site. The model of interface binding begins to explain the known specificity of lipoprotein-bound substrates and how the active site can be both close to the hydrophobic-hydrophilic interface and at the same time be accessible to the aqueous phase.
Figures


Similar articles
-
Crystallization and preliminary X-ray crystallographic analysis of human plasma platelet activating factor acetylhydrolase.Protein Pept Lett. 2009;16(1):97-100. doi: 10.2174/092986609787049321. Protein Pept Lett. 2009. PMID: 19149681 Free PMC article.
-
Platelet-activating factor acetylhydrolases: broad substrate specificity and lipoprotein binding does not modulate the catalytic properties of the plasma enzyme.Biochemistry. 2001 Apr 17;40(15):4539-49. doi: 10.1021/bi002600g. Biochemistry. 2001. PMID: 11294621
-
Identification of a domain that mediates association of platelet-activating factor acetylhydrolase with high density lipoprotein.J Biol Chem. 2008 Jun 20;283(25):17099-106. doi: 10.1074/jbc.M802394200. Epub 2008 Apr 22. J Biol Chem. 2008. PMID: 18434304 Free PMC article.
-
Platelet-activating factor acetylhydrolase.Prostaglandins Other Lipid Mediat. 2002 Aug;68-69:83-94. doi: 10.1016/s0090-6980(02)00023-0. Prostaglandins Other Lipid Mediat. 2002. PMID: 12432911 Review.
-
Platelet activating-factor acetylhydrolase II: A member of phospholipase A2 family that hydrolyzes oxidized phospholipids.Chem Phys Lipids. 2021 Sep;239:105103. doi: 10.1016/j.chemphyslip.2021.105103. Epub 2021 Jun 8. Chem Phys Lipids. 2021. PMID: 34116047 Review.
Cited by
-
Molecular Model of Plasma PAF Acetylhydrolase-Lipoprotein Association: Insights from the Structure.Pharmaceuticals (Basel). 2010 Mar 8;3(3):541-557. doi: 10.3390/ph3030541. Pharmaceuticals (Basel). 2010. PMID: 27713267 Free PMC article. Review.
-
FunSAV: predicting the functional effect of single amino acid variants using a two-stage random forest model.PLoS One. 2012;7(8):e43847. doi: 10.1371/journal.pone.0043847. Epub 2012 Aug 24. PLoS One. 2012. PMID: 22937107 Free PMC article.
-
Human Secretary Phospholipase A2 Mutations and Their Clinical Implications.J Inflamm Res. 2020 Sep 16;13:551-561. doi: 10.2147/JIR.S269557. eCollection 2020. J Inflamm Res. 2020. PMID: 32982370 Free PMC article. Review.
-
Lipoprotein-associated phospholipase A(2) interacts with phospholipid vesicles via a surface-disposed hydrophobic α-helix.Biochemistry. 2011 Jun 14;50(23):5314-21. doi: 10.1021/bi101916w. Epub 2011 May 17. Biochemistry. 2011. PMID: 21553808 Free PMC article.
-
A robust all-atom model for LCAT generated by homology modeling.J Lipid Res. 2015 Mar;56(3):620-634. doi: 10.1194/jlr.M056382. Epub 2015 Jan 14. J Lipid Res. 2015. PMID: 25589508 Free PMC article.
References
-
- Prescott, S. M., Zimmerman, G. A., and McIntyre, T. M. (1990) J. Biol. Chem. 265 17381-17384 - PubMed
-
- Prescott, S. M., Zimmerman, G. A., Stafforini, D. M., and McIntyre, T. M. (2000) Annu. Rev. Biochem. 69 419-445 - PubMed
-
- Tjoelker, L. W., Wilder, C., Eberhardt, C., Stafforini, D. M., Dietsch, G., Schimpf, B., Hooper, S., Le Trong, H., Cousens, L. S., Zimmerman, G. A., Yamada, Y., McIntyre, T. M., Prescott, S. M., and Gray, P. W. (1995) Nature 374 549-553 - PubMed
-
- Schaloske, R. H., and Dennis, E. A. (2006) Biochim. Biophys. Acta 1761 1246-1259 - PubMed
-
- Stremler, K. E., Stafforini, D. M., Prescott, S. M., and McIntyre, T. M. (1991) J. Biol. Chem. 266 11095-11103 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
