

Protein kinase d regulates trafficking of dendritic membrane proteins in developing neurons
- PMID: 18784310
- PMCID: PMC2648138
- DOI: 10.1523/JNEUROSCI.1879-08.2008
Protein kinase d regulates trafficking of dendritic membrane proteins in developing neurons
Abstract
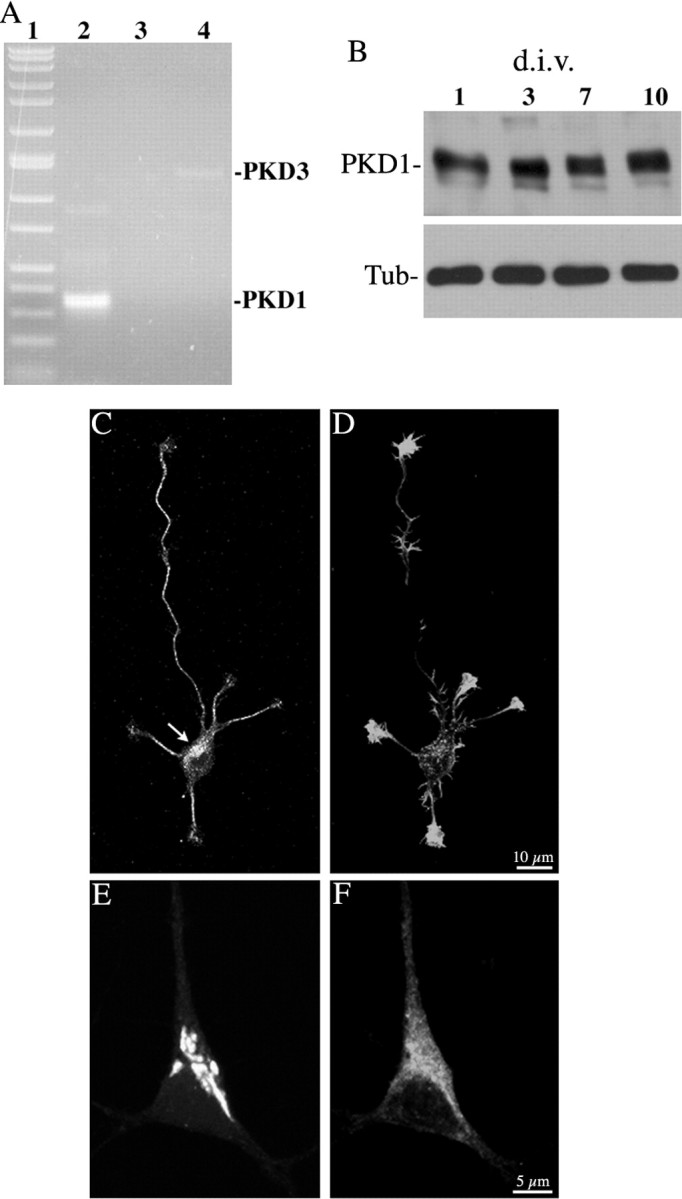
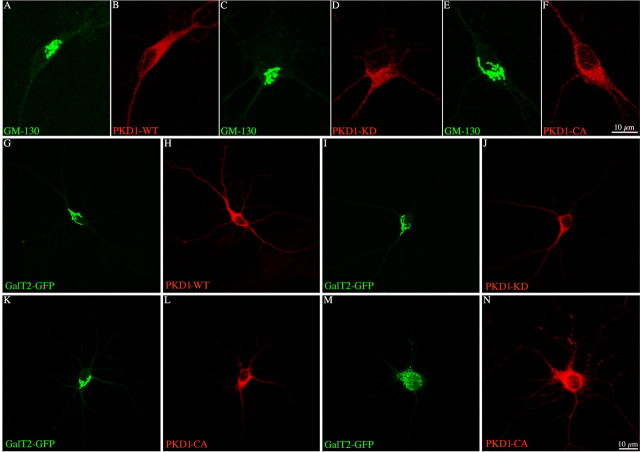
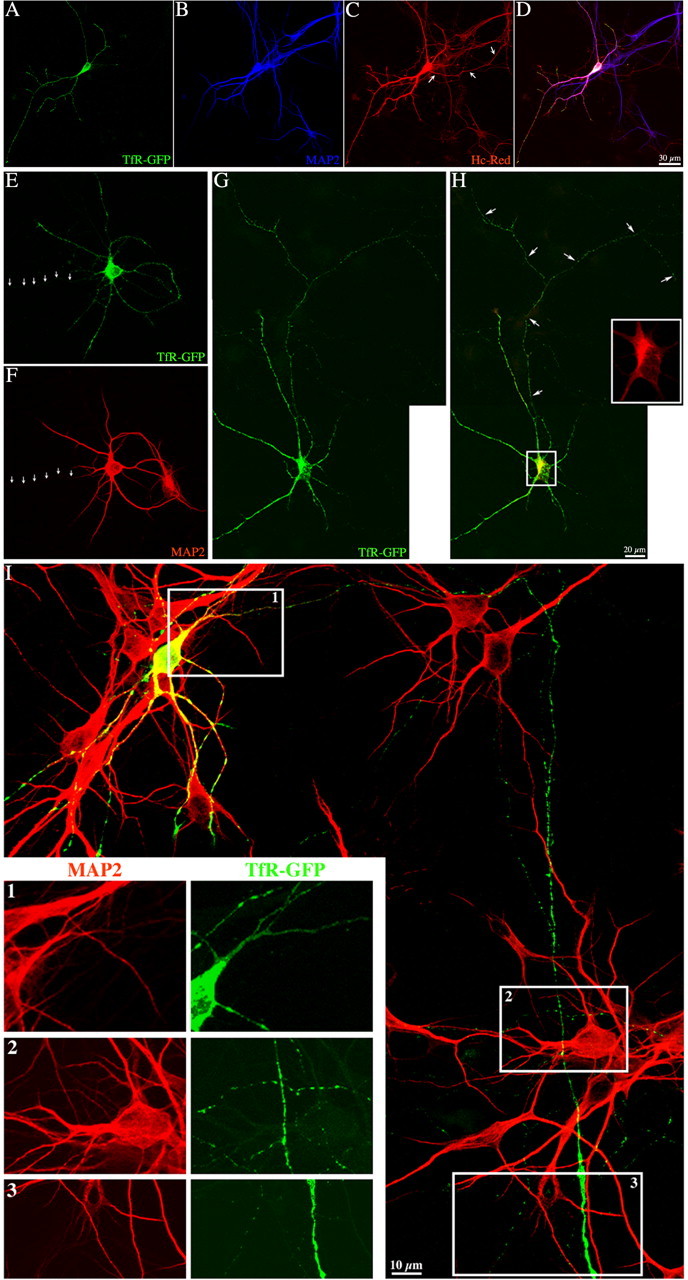
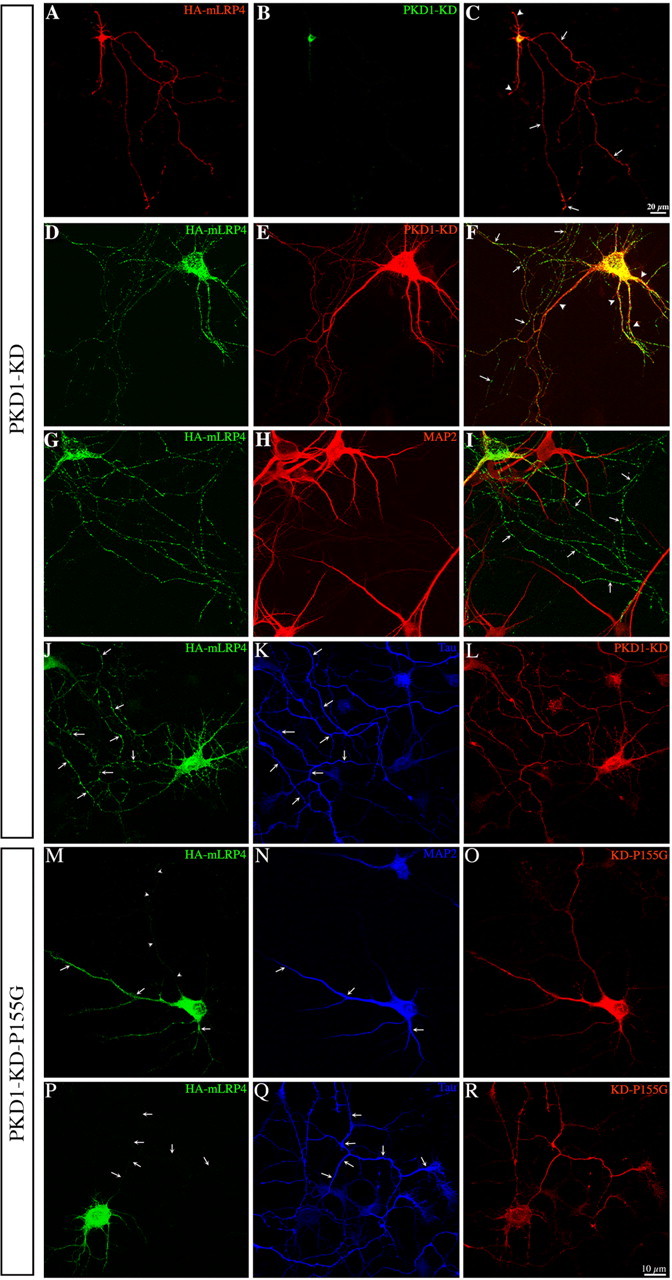
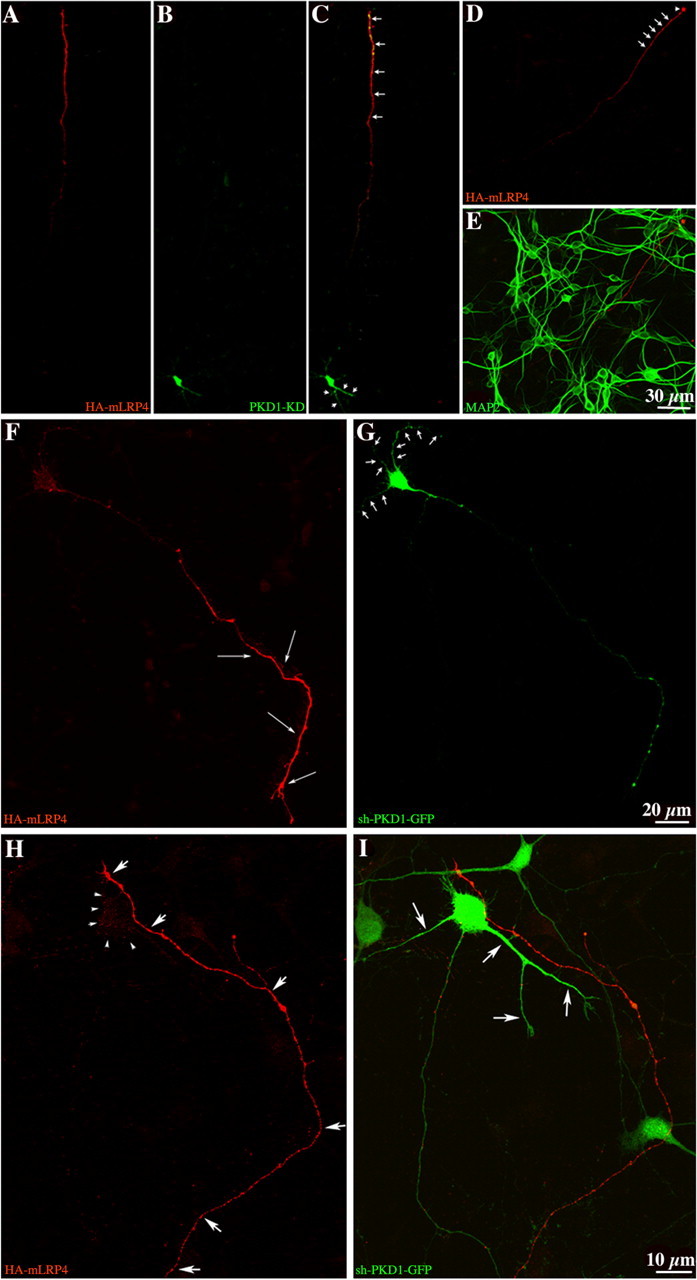
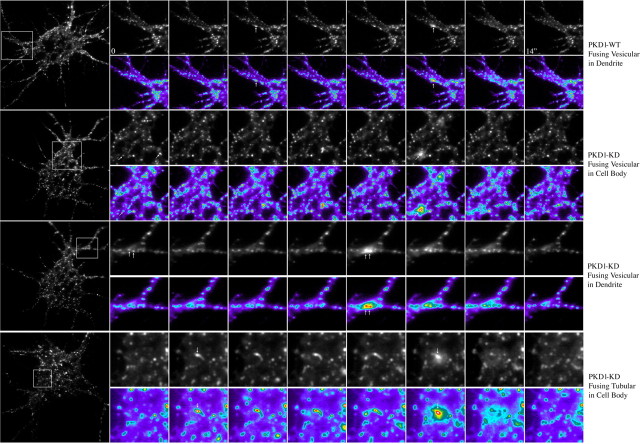
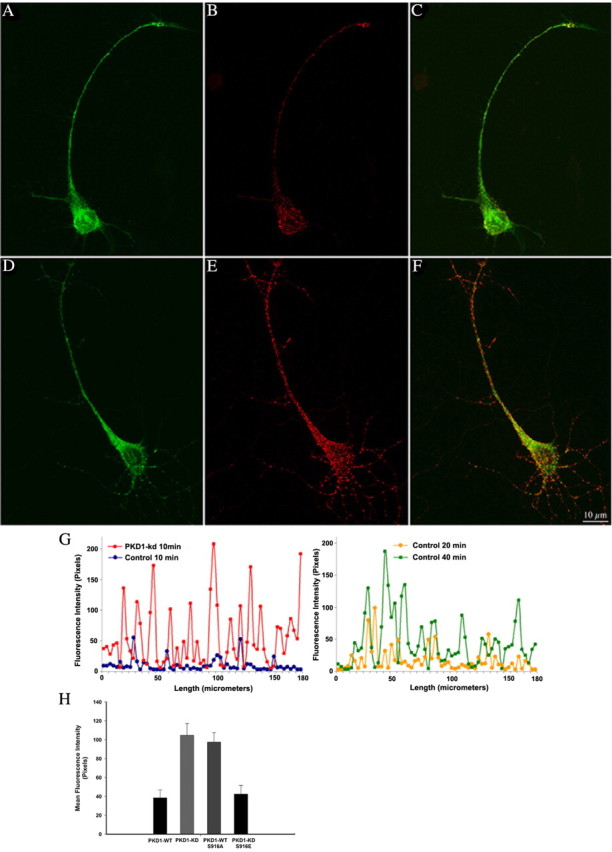
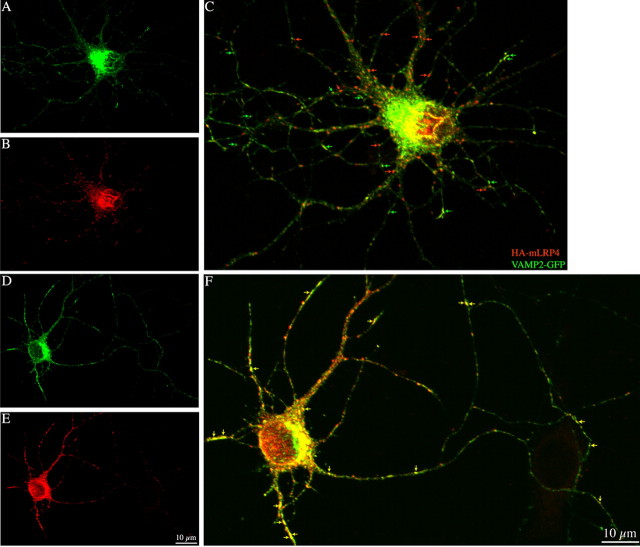
In non-neuronal cells, inactivation of protein kinase D (PKD) blocks fission of trans-Golgi network (TGN) transport carriers, inducing the appearance of long tubules filled with cargo. We now report on the function of PKD1 in neuronal protein trafficking. In cultured hippocampal pyramidal cells, the transferrin receptor (TfR) and the low-density receptor-related protein (LRP) are predominantly transported to dendrites and excluded from axons. Expression of kinase-inactive PKD1 or its depletion by RNA interference treatment dramatically and selectively alter the intracellular trafficking and membrane delivery of TfR- and LRP-containing vesicles, without inhibiting exit from the TGN or inducing Golgi tubulation. After PKD1 suppression, dendritic membrane proteins are mispackaged into carriers that transport VAMP2; these vesicles are distributed to both axons and dendrites, but are rapidly endocytosed from dendrites and preferentially delivered to the axonal membrane. A kinase-defective mutant of PKD1 lacking the ability to bind diacylglycerol and hence its Golgi localization does not cause missorting of TfR or LRP. These results suggest that in neurons PKD1 regulates TGN-derived sorting of dendritic proteins and hence has a role in neuronal polarity.
Figures
Similar articles
-
Protein kinase D controls the integrity of Golgi apparatus and the maintenance of dendritic arborization in hippocampal neurons.Mol Biol Cell. 2009 Apr;20(7):2108-20. doi: 10.1091/mbc.e08-09-0957. Epub 2009 Feb 11. Mol Biol Cell. 2009. PMID: 19211839 Free PMC article.
-
Both the establishment and maintenance of neuronal polarity require the activity of protein kinase D in the Golgi apparatus.J Neurosci. 2008 Aug 27;28(35):8832-43. doi: 10.1523/JNEUROSCI.1291-08.2008. J Neurosci. 2008. PMID: 18753385 Free PMC article.
-
Sorting and directed transport of membrane proteins during development of hippocampal neurons in culture.Proc Natl Acad Sci U S A. 2001 Jun 19;98(13):7051-7. doi: 10.1073/pnas.111146198. Proc Natl Acad Sci U S A. 2001. PMID: 11416186 Free PMC article.
-
Rapid dendritic transport of TGN38, a putative cargo receptor.Brain Res Mol Brain Res. 2004 Aug 23;127(1-2):68-78. doi: 10.1016/j.molbrainres.2004.05.013. Brain Res Mol Brain Res. 2004. PMID: 15306122
-
The role of selective transport in neuronal protein sorting.Neuron. 2000 May;26(2):465-72. doi: 10.1016/s0896-6273(00)81178-2. Neuron. 2000. PMID: 10839364
Cited by
-
The Golgi Apparatus: A Voyage through Time, Structure, Function and Implication in Neurodegenerative Disorders.Cells. 2023 Jul 31;12(15):1972. doi: 10.3390/cells12151972. Cells. 2023. PMID: 37566051 Free PMC article. Review.
-
Transcriptional regulation of neuronal polarity and morphogenesis in the mammalian brain.Neuron. 2011 Oct 6;72(1):22-40. doi: 10.1016/j.neuron.2011.09.018. Neuron. 2011. PMID: 21982366 Free PMC article. Review.
-
Stress-induced vulnerability to develop cocaine addiction depends on cofilin modulation.Neurobiol Stress. 2021 Jun 3;15:100349. doi: 10.1016/j.ynstr.2021.100349. eCollection 2021 Nov. Neurobiol Stress. 2021. PMID: 34169122 Free PMC article.
-
Protein kinase D controls the integrity of Golgi apparatus and the maintenance of dendritic arborization in hippocampal neurons.Mol Biol Cell. 2009 Apr;20(7):2108-20. doi: 10.1091/mbc.e08-09-0957. Epub 2009 Feb 11. Mol Biol Cell. 2009. PMID: 19211839 Free PMC article.
-
Identification of a de novo mutation of the FOXG1 gene and comprehensive analysis for molecular factors in Chinese FOXG1-related encephalopathies.Front Mol Neurosci. 2022 Dec 7;15:1039990. doi: 10.3389/fnmol.2022.1039990. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36568277 Free PMC article.
References
-
- Baron CL, Malhotra V. Role of diacylglycerol in PKD recruitment to the TGN and protein transport to the plasma membrane. Science. 2002;295:325–328. - PubMed
-
- Bowden ET, Barth M, Thomas D, Glazer RI, Mueller SC. An invasion-related complex of cortactin, paxillin and PKCmu associates with invadopodia at sites of extracellular matrix degradation. Oncogene. 1999;18:4440–4449. - PubMed
-
- Burack MA, Silverman MA, Banker G. The role of selective transport in neuronal polarity sorting. Neuron. 2000;26:465–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous