

Review
doi: 10.1016/j.cub.2008.07.001.
Temporal organization of the cell cycle
Affiliations
- PMID: 18786381
- PMCID: PMC2856080
- DOI: 10.1016/j.cub.2008.07.001
Item in Clipboard
Review
Temporal organization of the cell cycle
Curr Biol.
.
Abstract
The coordination of growth, DNA replication and division in proliferating cells can be adequately explained by a 'clock + checkpoint' model. The clock is an underlying cyclical sequence of states; the checkpoints ensure that the cycle proceeds without mistakes. From the molecular complexities of the control system in modern eukaryotes, we isolate a simple network of positive and negative feedbacks that embodies a 'clock + checkpoints'. The model accounts for the fundamental physiological properties of mitotic cell divisions, evokes a new view of the meiotic program, and suggests how the control system may have evolved in the first place.
Figures

At the bottom-left of the diagram cell division gives rise to two small daughter cells. The nucleus of each cell contains two copies of each chromosome (‘diploidy’): one from mother (red) and one from father (blue). Each chromosome contains a single double-stranded DNA molecule. During S phase (DNA synthesis), every DNA molecule is replicated. When this process is completed, the cell is in G2 phase (the two sister chromatids are held together by cohesion complexes, yellow). From G2 phase, the cell enters mitosis (M phase), which consists of four subphases: Prophase (not shown; nuclear envelope breakdown and formation of mitotic spindle), Metaphase (all chromosomes aligned at the middle of the spindle, with sister chromatids attached to opposite poles of the spindle), Anaphase (not shown; dissolution of cohesins and separation of the sister chromatids to opposite sides of the cell), Telophase (reformation of nuclear envelopes around the two complete sets of unreplicated chromosomes). At cell division, the binucleate cell is constricted in the middle to give rise to two small, uninucleate daughter cells. In the center of the diagram, we indicate that major events of the cell cycle are triggered by a family of protein kinases called ‘cyclin-dependent kinases’. For example, Cdk1 in complex with cyclin B phosphorylates target proteins (blue) that drive progression through prophase into metaphase. To exit mitosis (anaphase, telophase, cell division), CDK activity must be removed and the target proteins must be returned to their unphosphorylated state by exit-specific phosphatases.
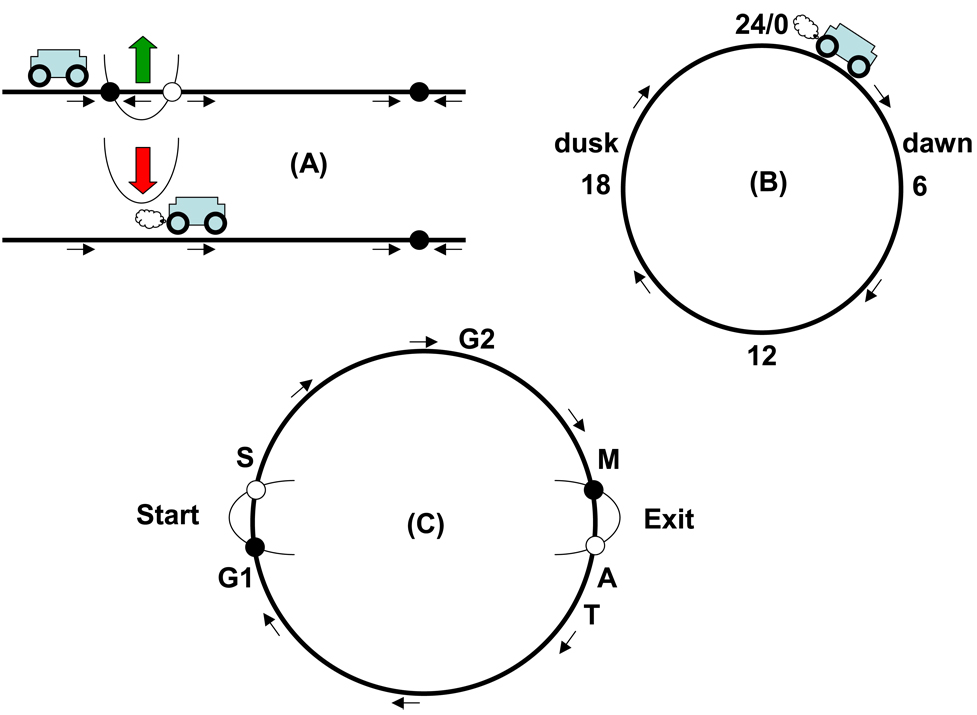
(A) A bistable switch creates a checkpoint. (B) A simple clock is a circular sequence of states. (C) A circular track with checkpoints models cell cycle progression. See text for details.
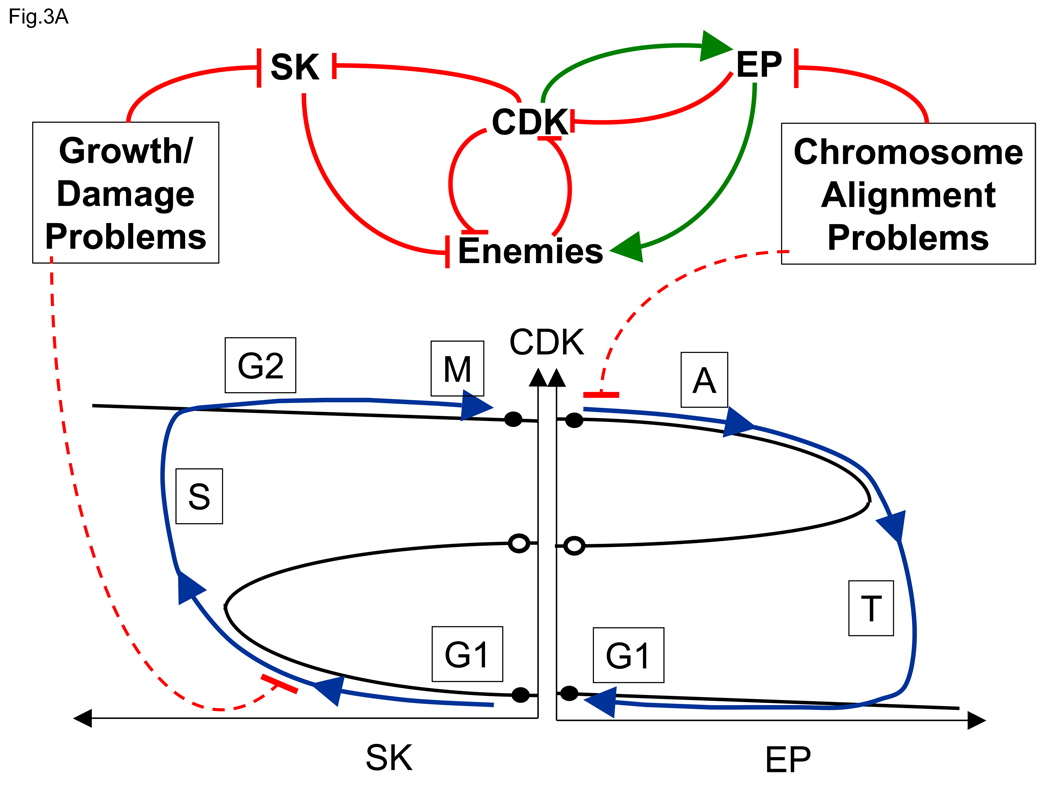
(A) CDK and its Enemies are involved in an antagonistic (double negative) feedback loop (top center), which can persist in either of two stable steady states: a G1-like state with low CDK activity and prominent Enemy forces, and an M-like steady state with high CDK activity and Enemies in retreat. These two stable steady states are represented by black circles along the CDK axis (bottom center). The pairs of circles side-by-side are meant to represent the same steady state under conditions when both SK (left) and EP (right) are close to zero. As in Figure 2(A), the two stable steady states are separated by an unstable steady state (open circle). A newborn cell in G1 (lower left) can be induced to enter S/G2/M by activation of a Starter Kinase (SK) that phosphorylates and weakens the Enemy forces, allowing CDK activity to rise and trigger S phase. Among other duties, CDK down-regulates SK, but the bistable switch remains in the upper state. Notice that, as SK activity increases, the stable G1-like steady state merges with and is annihilated by the unstable steady state, exactly as in Figure 2(A). Exit from mitosis (lower right) is induced by Exit Proteins (EP), which activate the proteins that destroy CDK activity. When the upper steady state merges with the unstable state state, the checkpoint is lifted and the cell returns to the G1 state. Because EP activity depends on CDK, after CDK falls, EP activity also disappears, but now the Enemies have the upper hand. The checkpoint barriers are responsive to two classes of problems. Growth requirements and damage repair processes (upper left) typically halt the cell cycle in G1 phase by preventing up-regulation of SK activity. Misalignment of chromosomes on the spindle (upper right) blocks exit from mitosis by preventing activation of EP. (B) Progression around the hysteresis loop in panel A corresponds to periodic changes in activities of CDK, Enemies, SK and EP. At Start , a burst of SK activity flips the CDK switch on , and at Exit , a burst of EP activity flips the CDK switch off .

(A) CDK and its Enemies are involved in an antagonistic (double negative) feedback loop (top center), which can persist in either of two stable steady states: a G1-like state with low CDK activity and prominent Enemy forces, and an M-like steady state with high CDK activity and Enemies in retreat. These two stable steady states are represented by black circles along the CDK axis (bottom center). The pairs of circles side-by-side are meant to represent the same steady state under conditions when both SK (left) and EP (right) are close to zero. As in Figure 2(A), the two stable steady states are separated by an unstable steady state (open circle). A newborn cell in G1 (lower left) can be induced to enter S/G2/M by activation of a Starter Kinase (SK) that phosphorylates and weakens the Enemy forces, allowing CDK activity to rise and trigger S phase. Among other duties, CDK down-regulates SK, but the bistable switch remains in the upper state. Notice that, as SK activity increases, the stable G1-like steady state merges with and is annihilated by the unstable steady state, exactly as in Figure 2(A). Exit from mitosis (lower right) is induced by Exit Proteins (EP), which activate the proteins that destroy CDK activity. When the upper steady state merges with the unstable state state, the checkpoint is lifted and the cell returns to the G1 state. Because EP activity depends on CDK, after CDK falls, EP activity also disappears, but now the Enemies have the upper hand. The checkpoint barriers are responsive to two classes of problems. Growth requirements and damage repair processes (upper left) typically halt the cell cycle in G1 phase by preventing up-regulation of SK activity. Misalignment of chromosomes on the spindle (upper right) blocks exit from mitosis by preventing activation of EP. (B) Progression around the hysteresis loop in panel A corresponds to periodic changes in activities of CDK, Enemies, SK and EP. At Start , a burst of SK activity flips the CDK switch on , and at Exit , a burst of EP activity flips the CDK switch off .

At meiosis I (far right), homologous chromosomes align on the spindle, with identical replicated chromosomes attached to opposite poles of the spindle. At anaphase of meiosis I, the arm-cohesins, which hold homologous chromosomes together are dissolved, but the centromeric cohesins, which hold sister chromatids together, remain intact. The replicated chromosomes are segregated to opposite sides of the cell, and cell division generates haploid nuclei with replicated chromosomes. These nuclei enter meiosis II, during which the sister chromatids are segregated, producing haploid gametes in G1 phase (unreplicated chromosomes).

(A) Steady state activity of CDK as a function of both SK and EP activities. The CDK surface is a pleated sheet. The upper surface of the pleat is the high-CDK state (G2/Meta/Ana) and the lower surface is the low-CDK state (G1) in Figure 3A, bottom. (B) Projection of the pleated surface onto the SK-EP plane. White: bistable region. Color: indicates CDK activity in the monostable region (blue = high, red = low, purple = intermediate.)
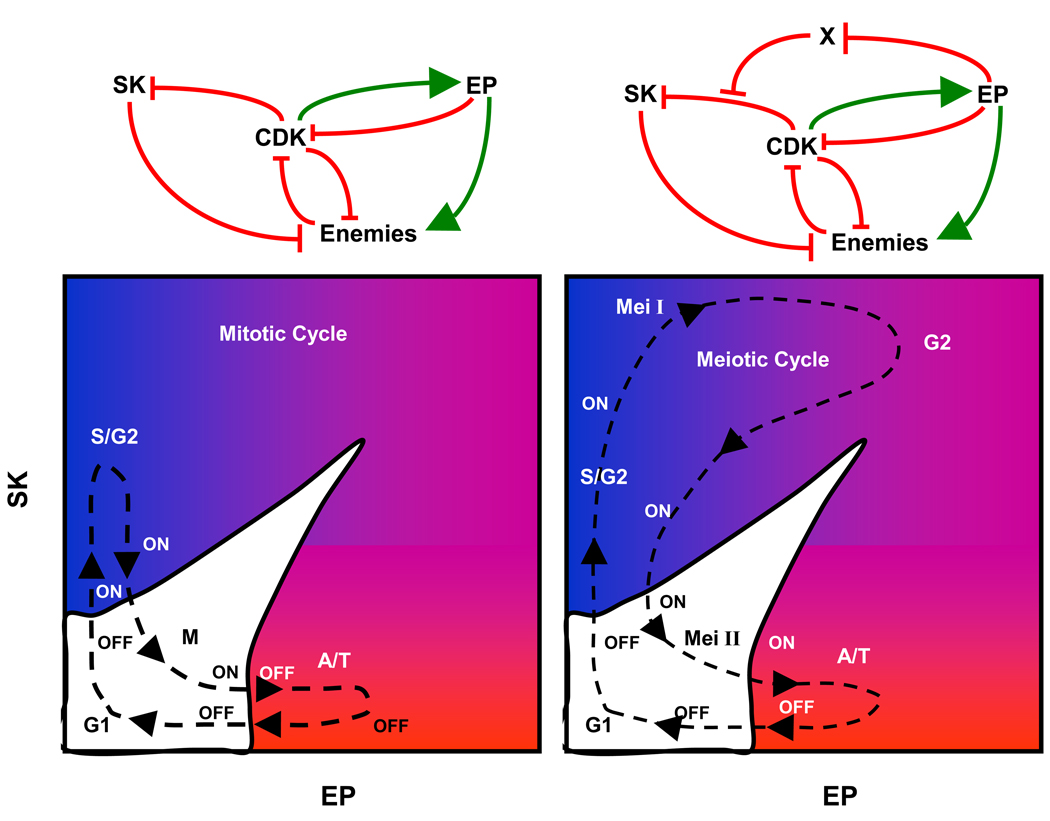
Left: Mitotic cell cycle. A newborn cell starts near the origin (SK ≈ 0, EP ≈ 0) on the lower surface (off ), with low CDK activity. First SK is activated (with EP ≈ 0), pushing the control system onto the upper surface, with high CDK activity. Then SK is removed, with EP activity still small. The cell halts on the upper sheet (on ) at metaphase, until EP can be activated. As EP rises (with SK ≈ 0), the control system is forced to jump to the lower surface, returning to G1 as EP disappears. Right: Meiosis I is a variant of mitosis, induced, we propose, by a single hypothetical protein, X, that blocks the down-regulation of SK by CDK. A limited supply of X is made during G1 phase of the first meiotic cycle, we suppose. S phase of meiosis I requires up-regulation of SK, exactly as in a mitotic cycle, except that SK cannot be down-regulated by CDK. The cell exits meiosis I by up-regulating EP with SK high. CDK activity does not fall low enough to permit relicensing of origins of DNA replication, so the cell enters a G2-like state of the control system after meiosis I. As EP destroys the limited supply of X, CDK can now remove SK. The cell enters meiosis II in much the same state as a normal mitotic division. Upon exiting meiosis II, the cell is now in G1 with un-replicated, haploid chromosomes.
References
-
- Mitchison JM. The Biology of the Cell Cycle. Cambridge UK: Cambridge Univ. Press; 1971.
-
- Morgan DO. Principles of CDK regulation. Nature. 1995;374:131–134. - PubMed
-
- Nurse P. A long twentieth century of the cell cycle and beyond. Cell. 2000;100:71–78. - PubMed
-
- Kelly TJ, Martin GS, Forsburg SL, Stephen RJ, Russo A, Nurse P. The fission yeast cdc18+ gene product couples S phase to START and mitosis. Cell. 1993;74:371–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
