

Noncanonical binding of calmodulin to aquaporin-0: implications for channel regulation
- PMID: 18786401
- PMCID: PMC2605016
- DOI: 10.1016/j.str.2008.06.011
Noncanonical binding of calmodulin to aquaporin-0: implications for channel regulation
Abstract
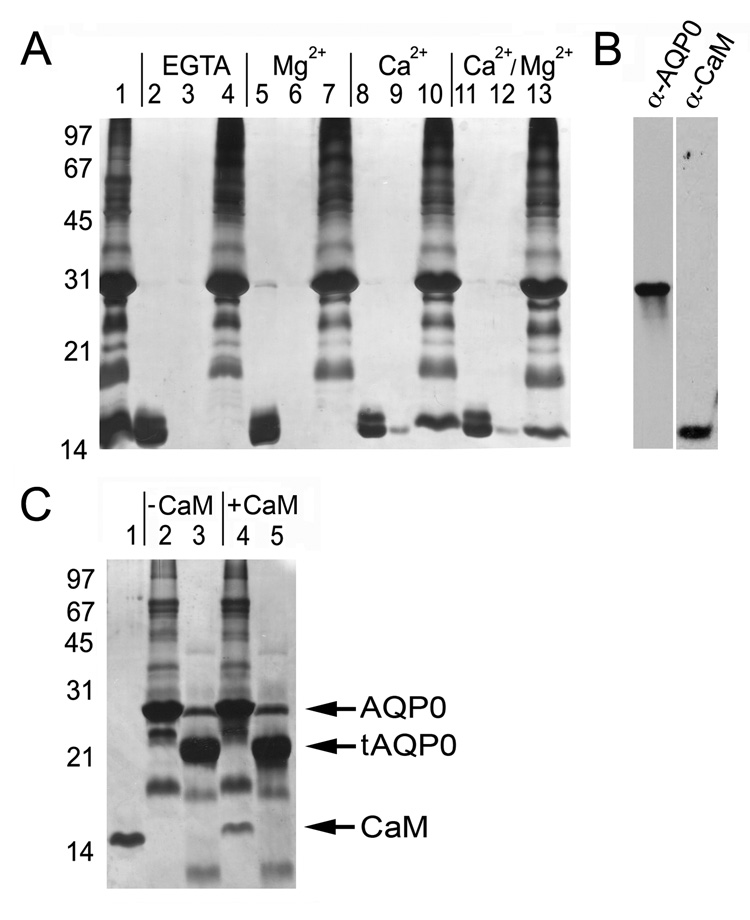
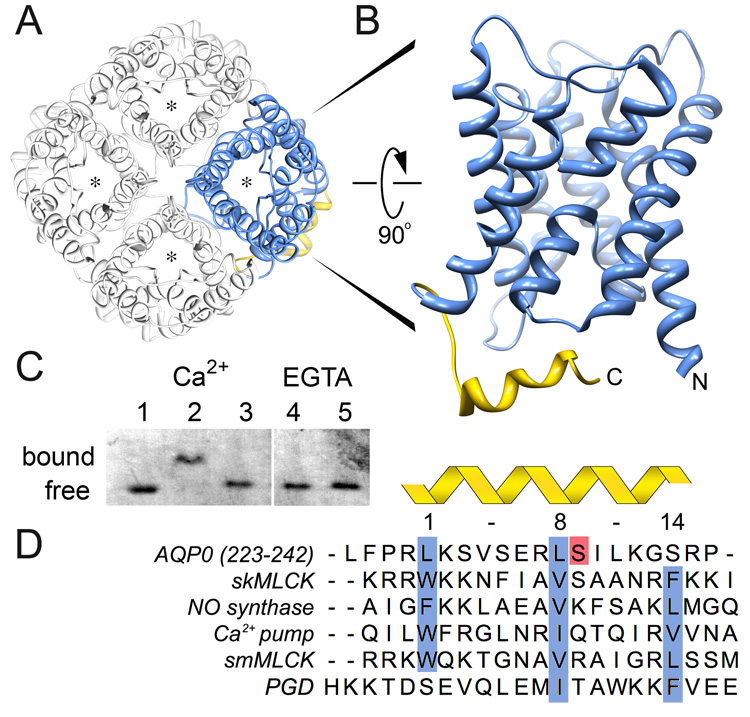
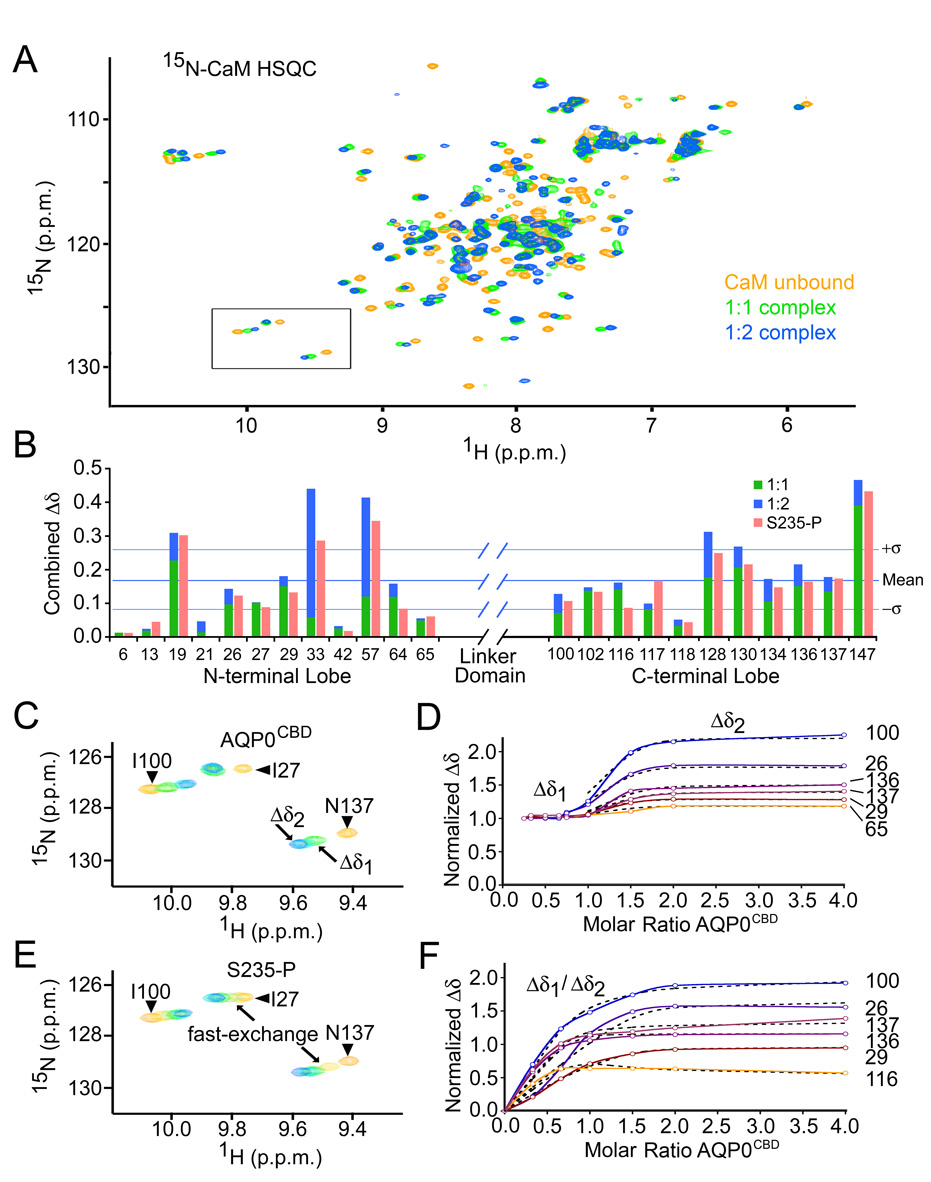
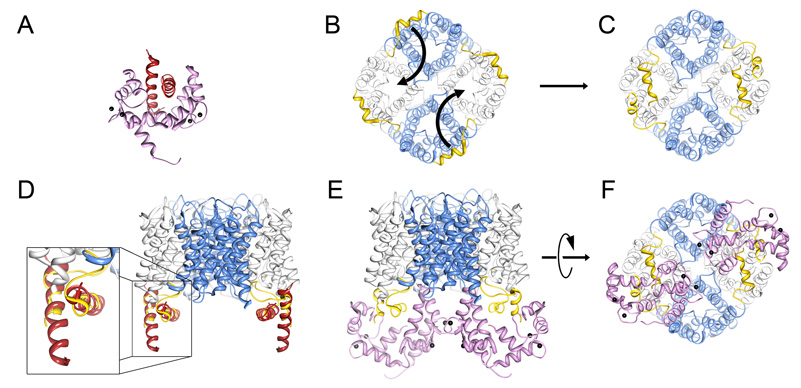
Aquaporins (AQPs) are a family of ubiquitous membrane channels that conduct water across cell membranes. AQPs form homotetramers containing four functional and independent water pores. Aquaporin-0 (AQP0) is expressed in the eye lens, where its water permeability is regulated by calmodulin (CaM). Here we use a combination of biochemical methods and NMR spectroscopy to probe the interaction between AQP0 and CaM. We show that CaM binds the AQP0 C-terminal domain in a calcium-dependent manner. We demonstrate that only two CaM molecules bind a single AQP0 tetramer in a noncanonical fashion, suggesting a form of cooperativity between AQP0 monomers. Based on these results, we derive a structural model of the AQP0/CaM complex, which suggests CaM may be inhibitory to channel permeability by capping the vestibules of two monomers within the AQP0 tetramer. Finally, phosphorylation within AQP0's CaM binding domain inhibits the AQP0/CaM interaction, suggesting a temporal regulatory mechanism for complex formation.
Figures

Similar articles
-
Calmodulin Gates Aquaporin 0 Permeability through a Positively Charged Cytoplasmic Loop.J Biol Chem. 2017 Jan 6;292(1):185-195. doi: 10.1074/jbc.M116.743724. Epub 2016 Sep 22. J Biol Chem. 2017. PMID: 27660387 Free PMC article.
-
Aquaporin 0-calmodulin interaction and the effect of aquaporin 0 phosphorylation.Biochemistry. 2008 Jan 8;47(1):339-47. doi: 10.1021/bi701980t. Epub 2007 Dec 15. Biochemistry. 2008. PMID: 18081321
-
Allosteric mechanism of water-channel gating by Ca2+-calmodulin.Nat Struct Mol Biol. 2013 Sep;20(9):1085-92. doi: 10.1038/nsmb.2630. Epub 2013 Jul 28. Nat Struct Mol Biol. 2013. PMID: 23893133 Free PMC article.
-
Junction-forming aquaporins.Curr Opin Struct Biol. 2008 Apr;18(2):229-35. doi: 10.1016/j.sbi.2007.11.003. Epub 2008 Jan 14. Curr Opin Struct Biol. 2008. PMID: 18194855 Free PMC article. Review.
-
Structural function of MIP/aquaporin 0 in the eye lens; genetic defects lead to congenital inherited cataracts.Handb Exp Pharmacol. 2009;(190):265-97. doi: 10.1007/978-3-540-79885-9_14. Handb Exp Pharmacol. 2009. PMID: 19096783 Review.
Cited by
-
Covalently Linking Oligomerization-Impaired GlpF Protomers Does Not Completely Re-establish Wild-Type Channel Activity.Int J Mol Sci. 2019 Feb 20;20(4):927. doi: 10.3390/ijms20040927. Int J Mol Sci. 2019. PMID: 30791644 Free PMC article.
-
Spatial analysis of human lens aquaporin-0 post-translational modifications by MALDI mass spectrometry tissue profiling.Exp Eye Res. 2011 Dec;93(6):912-20. doi: 10.1016/j.exer.2011.10.007. Epub 2011 Oct 25. Exp Eye Res. 2011. PMID: 22036630 Free PMC article.
-
Molecular mechanisms governing aquaporin relocalisation.Biochim Biophys Acta Biomembr. 2022 Apr 1;1864(4):183853. doi: 10.1016/j.bbamem.2021.183853. Epub 2021 Dec 30. Biochim Biophys Acta Biomembr. 2022. PMID: 34973181 Free PMC article. Review.
-
BFSP1 C-terminal domains released by post-translational processing events can alter significantly the calcium regulation of AQP0 water permeability.Exp Eye Res. 2019 Aug;185:107585. doi: 10.1016/j.exer.2019.02.001. Epub 2019 Feb 18. Exp Eye Res. 2019. PMID: 30790544 Free PMC article.
-
Evaluating protein complexes between human aquaporin and calmodulin using biomolecular fluorescence complementation.Sci Rep. 2025 Aug 2;15(1):28203. doi: 10.1038/s41598-025-12865-z. Sci Rep. 2025. PMID: 40750988 Free PMC article.
References
-
- Agre P, Kozono D. Aquaporin water channels: molecular mechanisms for human diseases. FEBS Lett. 2003;555:72–78. - PubMed
-
- Agre P, Sasaki S, Chrispeels MJ. Aquaporins: a family of water channel proteins. Am J Physiol. 1993;265:F461. - PubMed
-
- Alcala J, Lieska N, Maisel H. Protein composition of bovine lens cortical fiber cell membranes. Exp Eye Res. 1975;21:581–595. - PubMed
-
- Arneson ML, Cheng HL, Louis CF. Characterization of the ovine-lens plasma-membrane protein-kinase substrates. Eur J Biochem. 1995;234:670–679. - PubMed
-
- Babu YS, Bugg CE, Cook WJ. Structure of calmodulin refined at 2.2 A resolution. J Mol Biol. 1988;204:191–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
