

Bacteriophage lambda stabilization by auxiliary protein gpD: timing, location, and mechanism of attachment determined by cryo-EM
- PMID: 18786402
- PMCID: PMC2614288
- DOI: 10.1016/j.str.2008.05.016
Bacteriophage lambda stabilization by auxiliary protein gpD: timing, location, and mechanism of attachment determined by cryo-EM
Abstract
We report the cryo-EM structure of bacteriophage lambda and the mechanism for stabilizing the 20-A-thick capsid containing the dsDNA genome. The crystal structure of the HK97 bacteriophage capsid fits most of the T = 7 lambda particle density with only minor adjustment. A prominent surface feature at the 3-fold axes corresponds to the cementing protein gpD, which is necessary for stabilization of the capsid shell. Its position coincides with the location of the covalent cross-link formed in the docked HK97 crystal structure, suggesting an evolutionary replacement of this gene product in lambda by autocatalytic chemistry in HK97. The crystal structure of the trimeric gpD, in which the 14 N-terminal residues required for capsid binding are disordered, fits precisely into the corresponding EM density. The N-terminal residues of gpD are well ordered in the cryo-EM density, adding a strand to a beta-sheet formed by the capsid proteins and explaining the mechanism of particle stabilization.
Figures
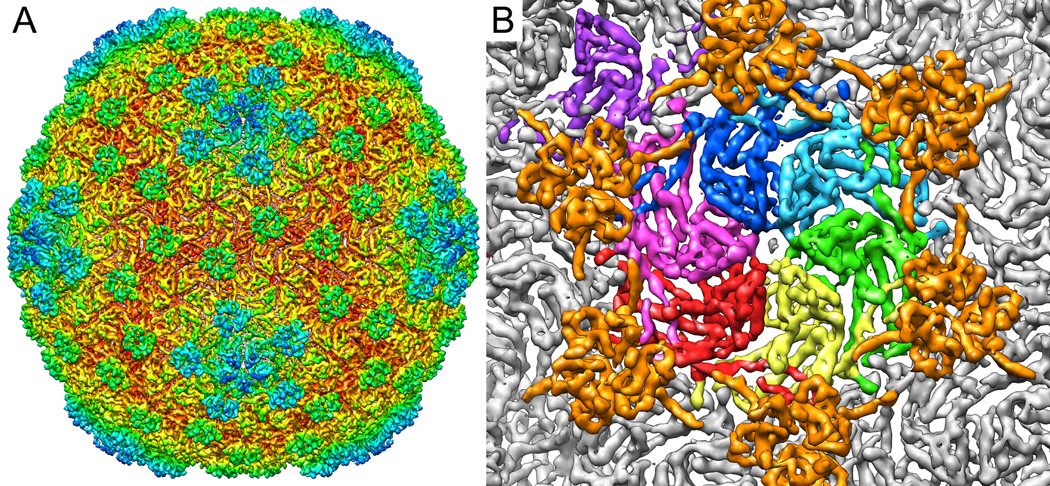
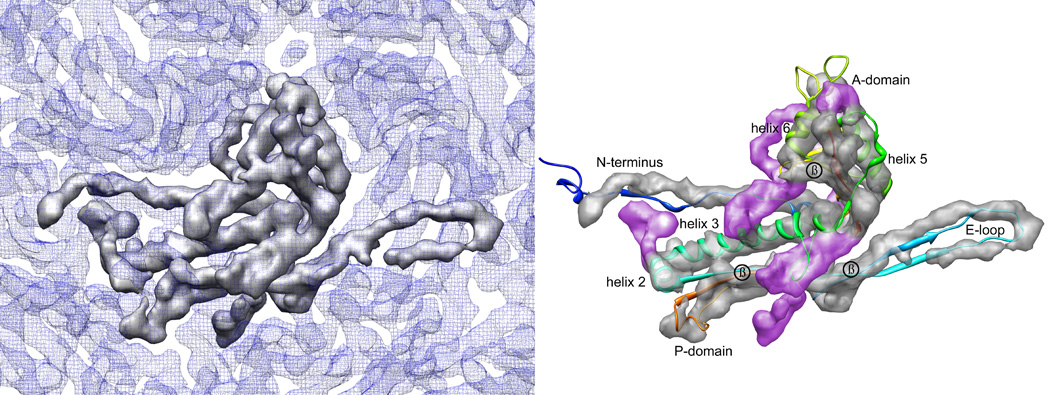
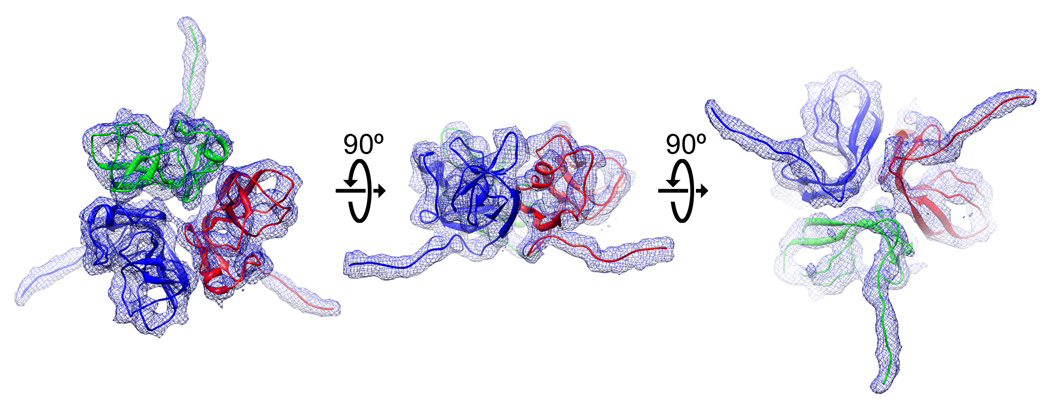
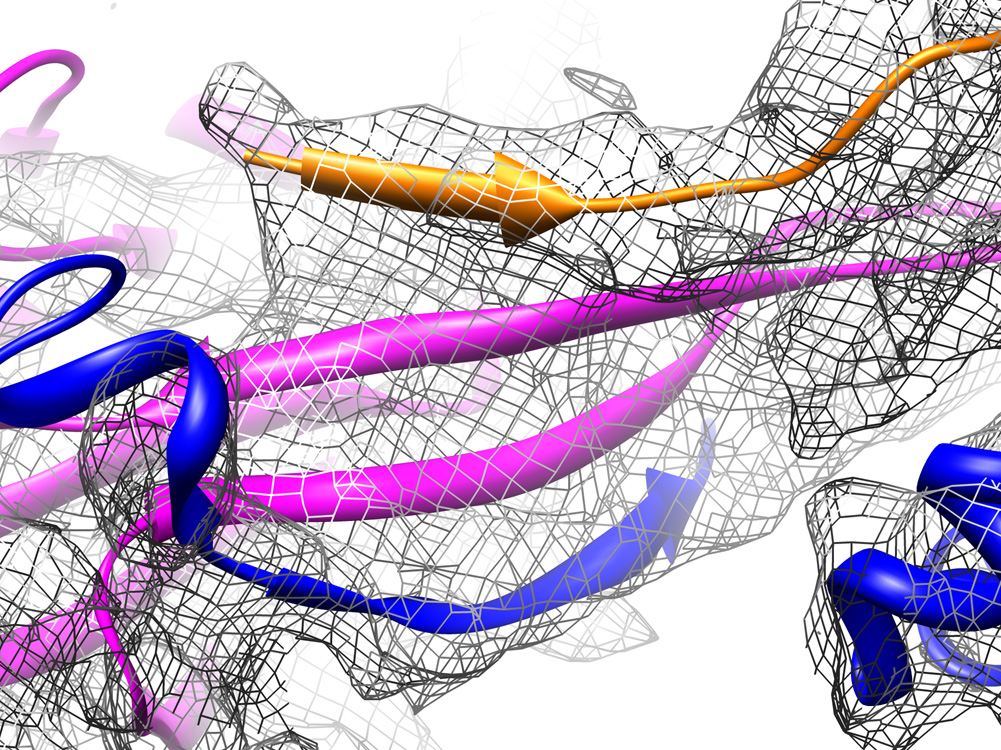
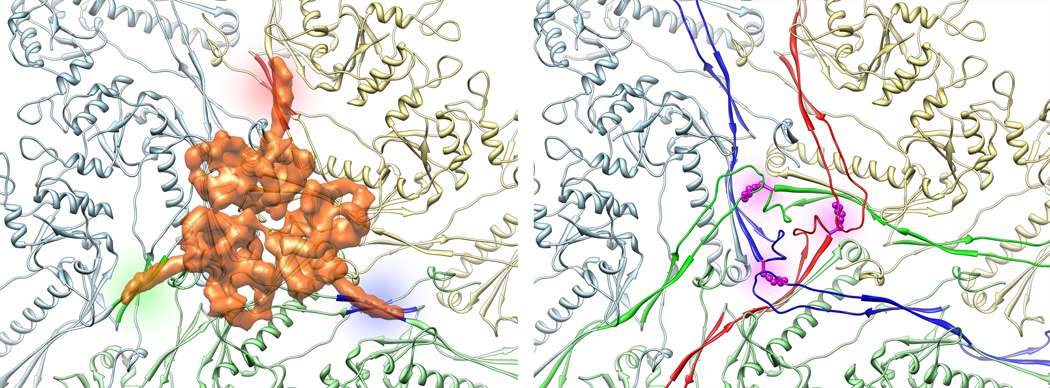
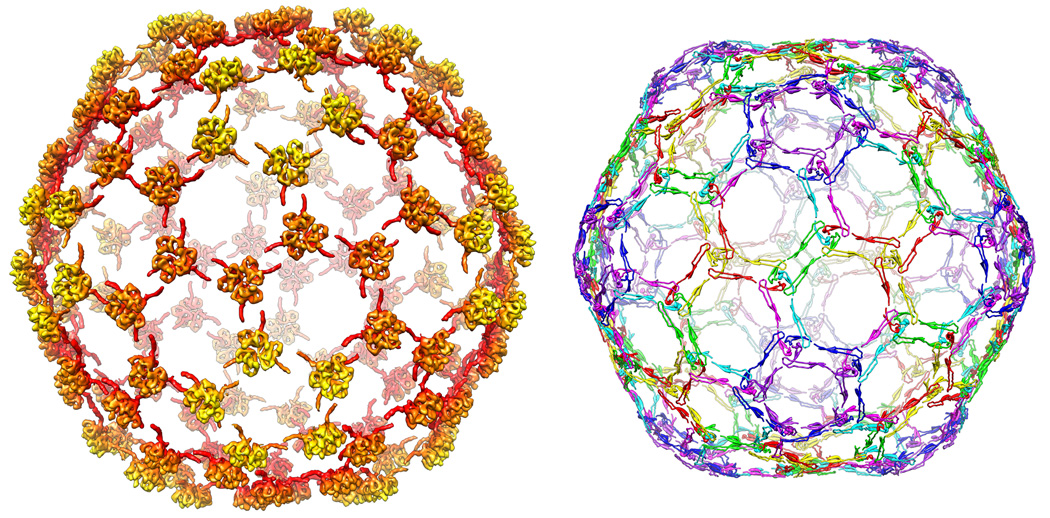
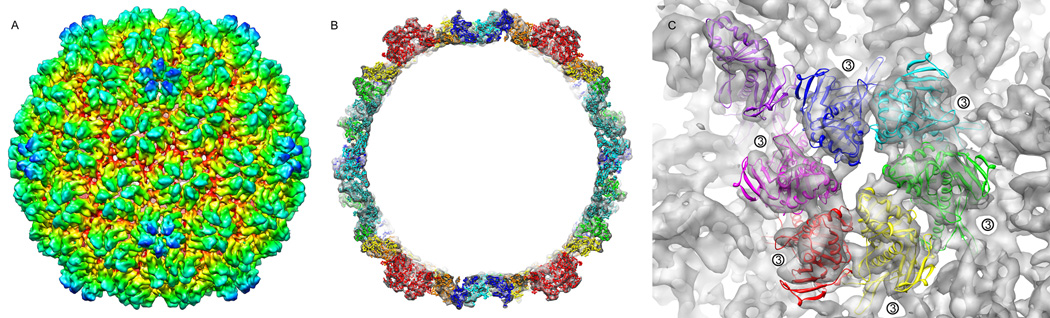
Comment in
-
Send for reinforcements! Conserved binding of capsid decoration proteins.Structure. 2008 Sep 10;16(9):1292-3. doi: 10.1016/j.str.2008.08.003. Structure. 2008. PMID: 18786391 No abstract available.
References
-
- Agirrezabala X, Velazquez-Muriel JA, Gomez-Puertas P, Scheres SH, Carazo JM, Carrascosa JL. Quasi-atomic model of bacteriophage t7 procapsid shell: insights into the structure and evolution of a basic fold. Structure. 2007;15:461–472. - PubMed
-
- Black LW. DNA packaging in dsDNA bacteriophages. Annu Rev Microbiol. 1989;43:267–292. - PubMed
-
- Campbell A. Comparative molecular biology of lambdoid phages. Annu Rev Microbiol. 1994;48:193–222. - PubMed
-
- Casjens SR. Comparative genomics and evolution of the tailed-bacteriophages. Curr Opin Microbiol. 2005;8:451–458. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
