

Deletion of CD151 results in a strain-dependent glomerular disease due to severe alterations of the glomerular basement membrane
- PMID: 18787104
- PMCID: PMC2543062
- DOI: 10.2353/ajpath.2008.071149
Deletion of CD151 results in a strain-dependent glomerular disease due to severe alterations of the glomerular basement membrane
Abstract
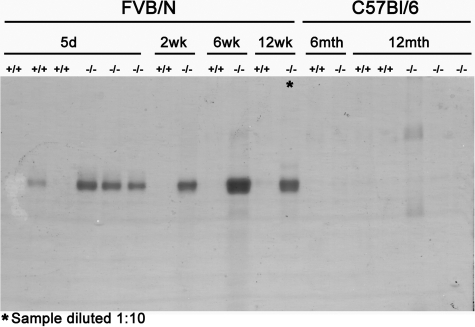
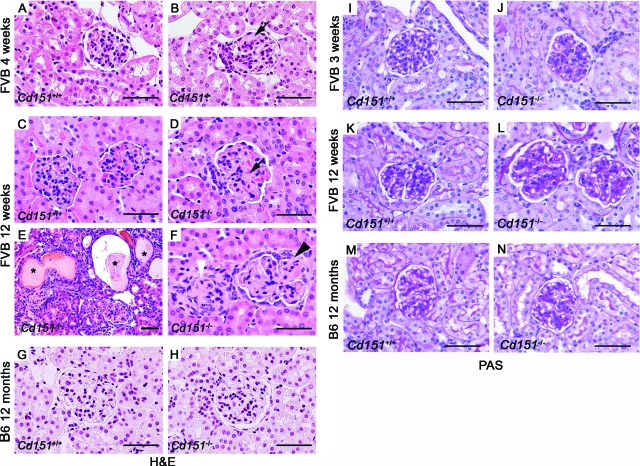
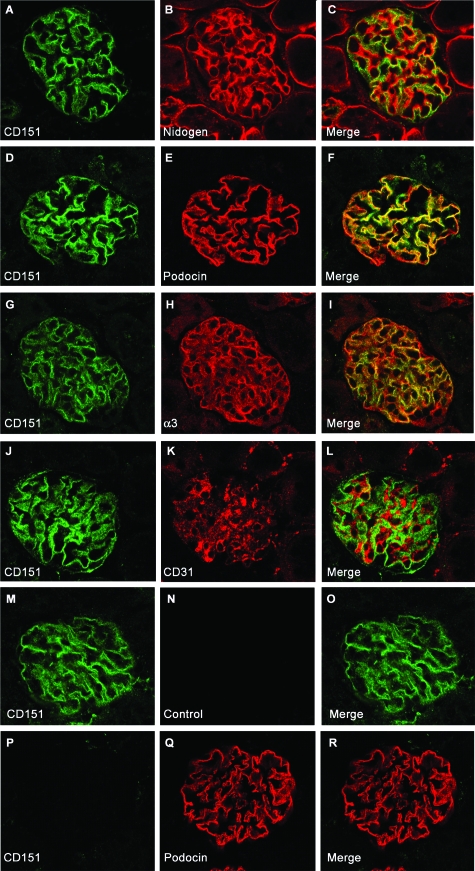
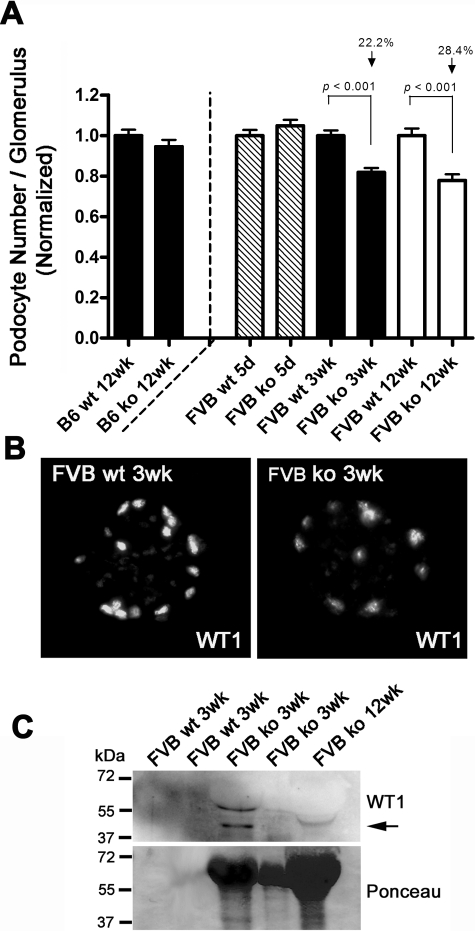
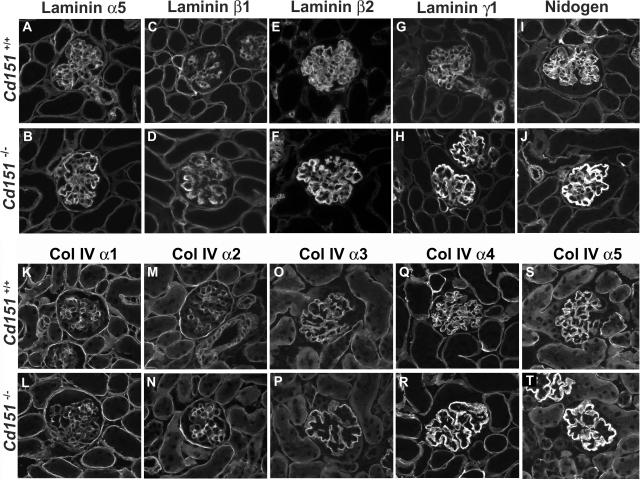
Alterations in CD151 have been associated with primary glomerular disease in both humans and mice, implicating CD151 as a key component of the glomerular filtration barrier. CD151 belongs to the tetraspanin family and associates with cell-matrix adhesion complexes such as alpha3beta1-integrin. Here we show that Cd151-deficient mice develop severe kidney disease on an FVB background but are healthy on a B6 background, providing a new and unique tool for the identification of genes that modulate the onset of proteinuria. To better understand the function of CD151 in the kidney, we studied its expression pattern and characterized early ultrastructural defects in Cd151-null kidneys. CD151 is expressed in podocytes of the mouse kidney and co-localizes with alpha3-integrin at the base of podocyte foot processes, at the site of anchorage to the glomerular basement membrane (GBM). Interestingly, the first ultrastructural lesions seen at the onset of proteinuria in Cd151-null kidneys were severe alterations of the GBM, reminiscent of Alport syndrome and consisting of massive thickening and splitting of the GBM. These lesions are associated with increased expression of GBM components. Podocyte abnormalities, effacement of foot processes, and podocyte loss appear to occur consequently to the GBM damage. In conclusion, CD151 appears to be involved in the establishment, maturation, and/or maintenance of the GBM structure in addition to its role in integrin-mediated adhesion strengthening.
Figures


References
-
- Johnstone DB, Holzman LB. Clinical impact of research on the podocyte slit diaphragm. Nat Clin Pract Nephrol. 2006;2:271–282. - PubMed
-
- Hudson BG, Reeders ST, Tryggvason K. Type IV collagen: structure, gene organization, and role in human diseases. Molecular basis of Goodpasture and Alport syndromes and diffuse leiomyomatosis. J Biol Chem. 1993;268:26033–26036. - PubMed
-
- Antignac C. Molecular genetics of basement membranes: the paradigm of Alport syndrome. Kidney Int Suppl. 1995;49:S29–S33. - PubMed
-
- Zenker M, Aigner T, Wendler O, Tralau T, Muntefering H, Fenski R, Pitz S, Schumacher V, Royer-Pokora B, Wuhl E, Cochat P, Bouvier R, Kraus C, Mark K, Madlon H, Dotsch J, Rascher W, Maruniak-Chudek I, Lennert T, Neumann LM, Reis A. Human laminin beta2 deficiency causes congenital nephrosis with mesangial sclerosis and distinct eye abnormalities. Hum Mol Genet. 2004;13:2625–2632. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
