

Late ROS accumulation and radiosensitivity in SOD1-overexpressing human glioma cells
- PMID: 18790046
- PMCID: PMC2637374
- DOI: 10.1016/j.freeradbiomed.2008.08.009
Late ROS accumulation and radiosensitivity in SOD1-overexpressing human glioma cells
Abstract
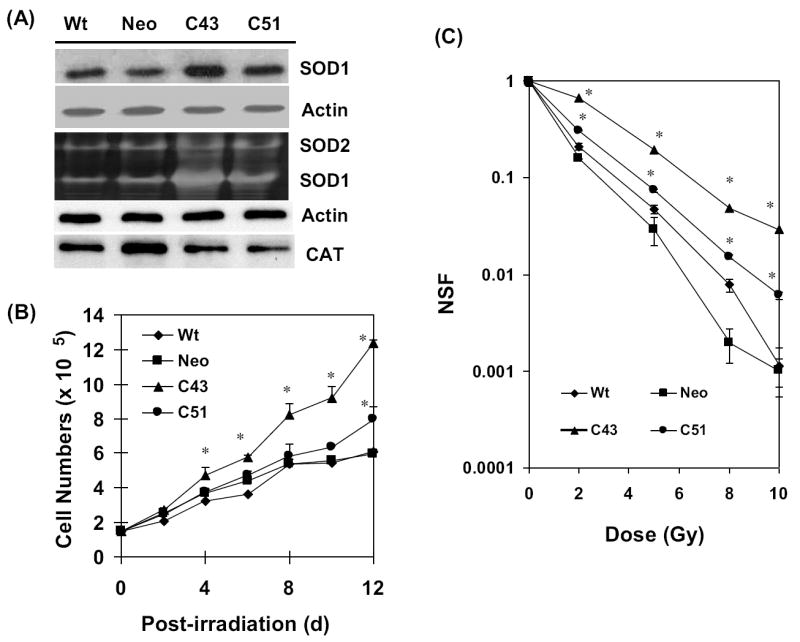
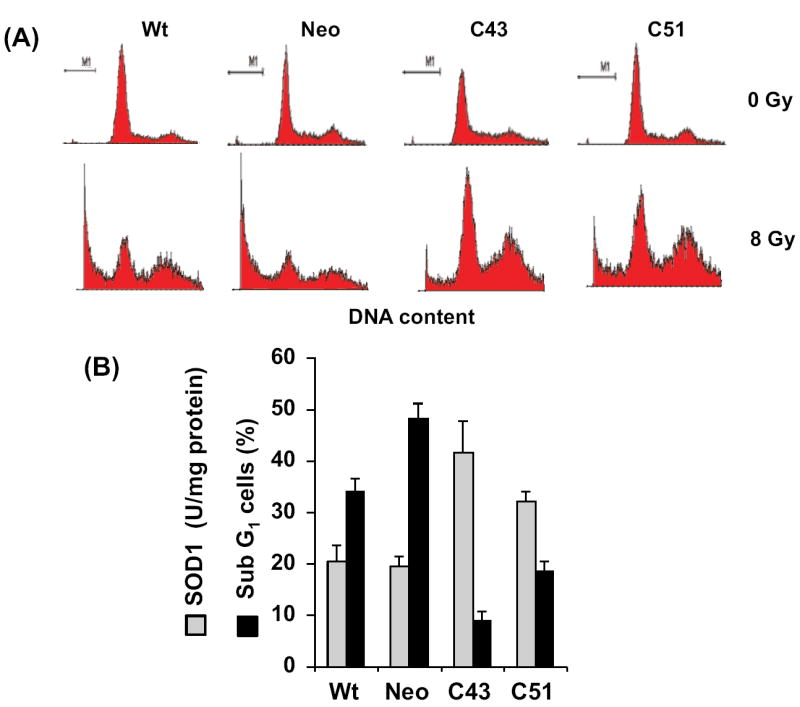
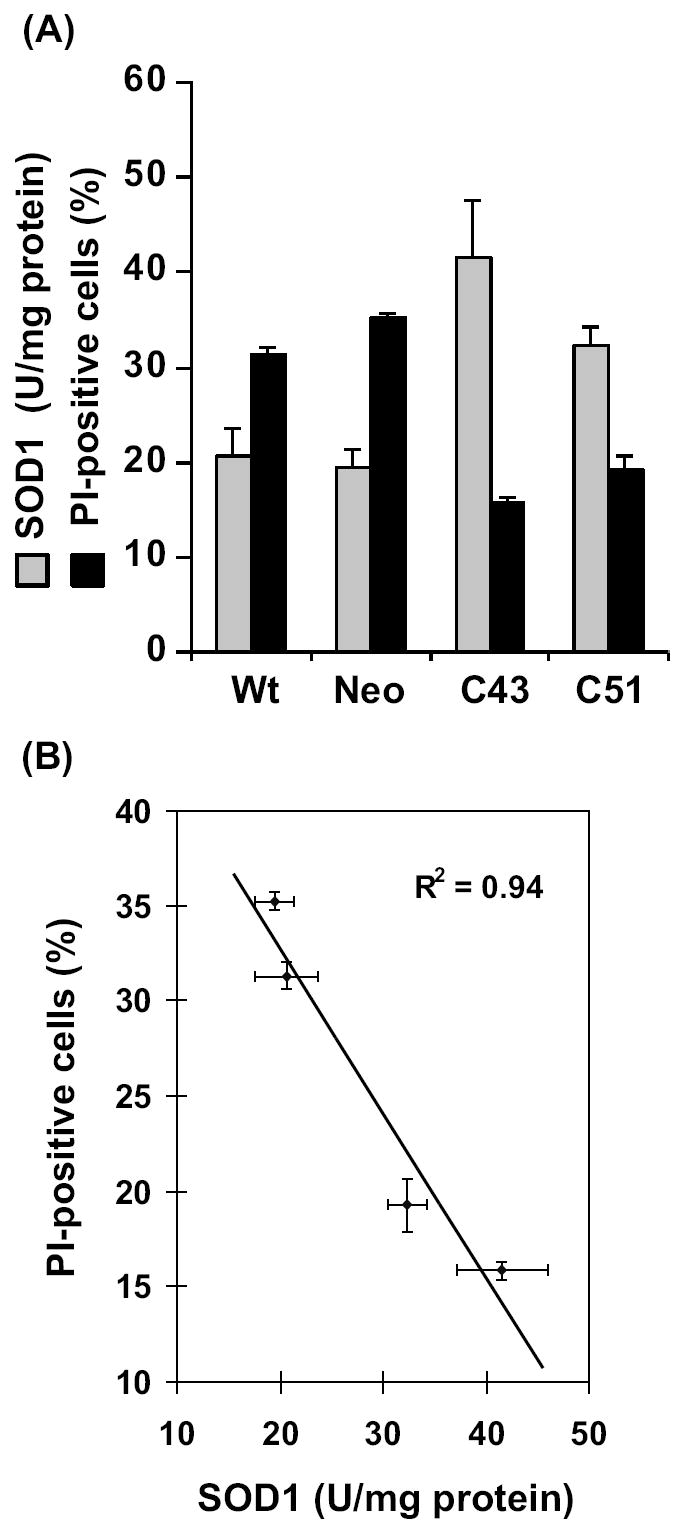
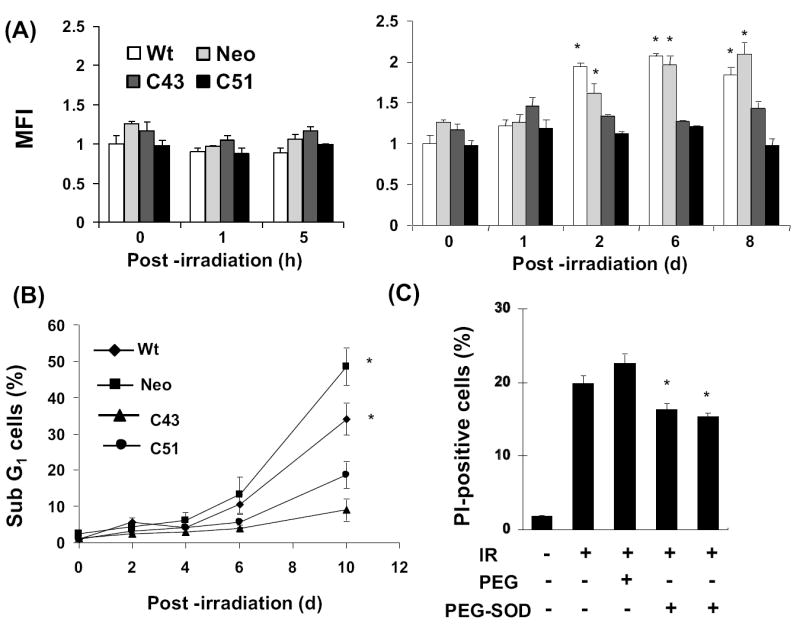
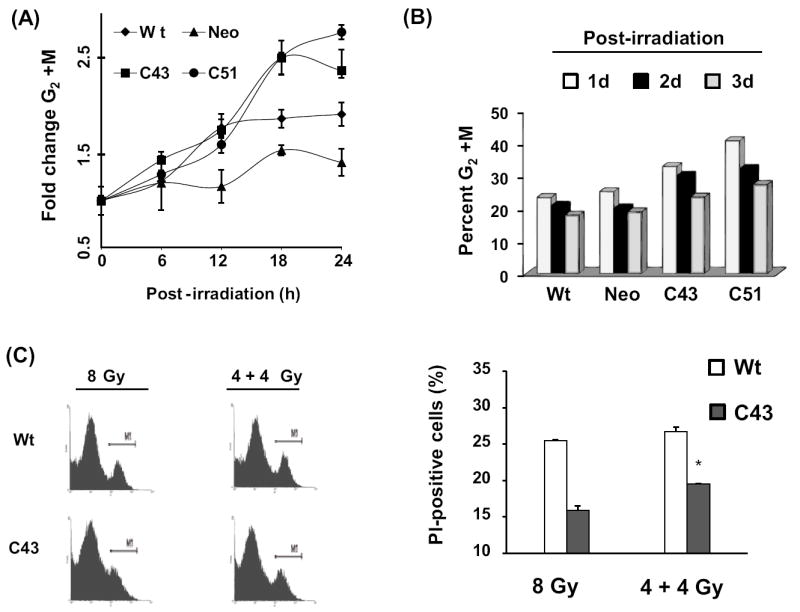
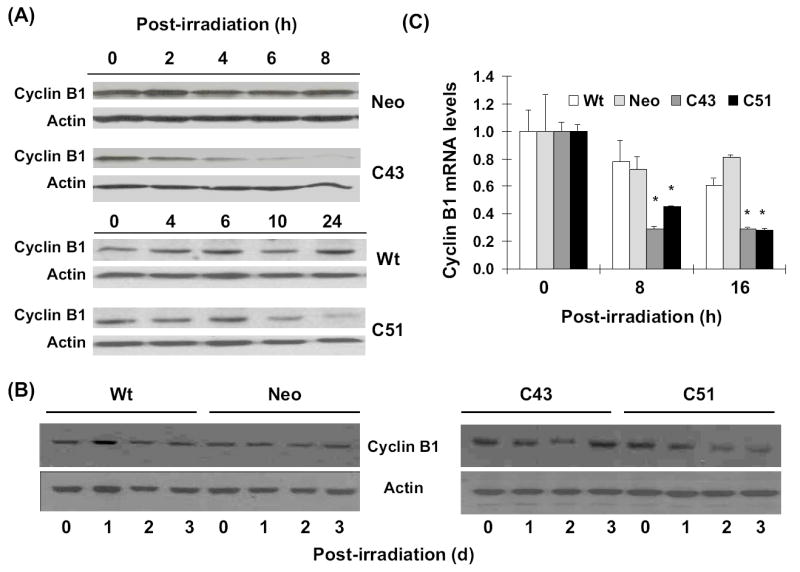
This study investigates the hypothesis that CuZn superoxide dismutase (SOD1) overexpression confers radioresistance to human glioma cells by regulating the late accumulation of reactive oxygen species (ROS) and the G(2)/M-checkpoint pathway. U118-9 human glioma cells (wild type, neo vector control, and stably overexpressing SOD1) were irradiated (0-10 Gy) and assayed for cell survival, cellular ROS levels, cell-cycle-phase distributions, and cyclin B1 expression. SOD1-overexpressing cells were radioresistant compared to wild-type (wt) and neo vector control (neo) cells. Irradiated wt and neo cells showed a significant increase (approximately twofold) in DHE fluorescence beginning at 2 days postirradiation, which remained elevated at 8 days postirradiation. Interestingly, the late accumulation of ROS was suppressed in irradiated SOD1-overexpressing cells. The increase in ROS levels was followed by a decrease in cell growth and viability and an increase in the percentage of cells with sub-G(1) DNA content. SOD1 overexpression enhanced radiation-induced G(2) accumulation within 24 h postirradiation, which was accompanied by a decrease in cyclin B1 mRNA and protein levels. These results support the hypothesis that long after radiation exposure a "metabolic redox response" regulates radiosensitivity of human glioma cells.
Figures
References
-
- Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006;160:1–40. - PubMed
-
- Zelko IN, Mariani TJ, Folz RJ. Superoxide dismutase multigene family: a comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic Biol Med. 2002;33:337–349. - PubMed
-
- Elchuri S, Oberley TD, Qi W, Eisenstein RS, Jackson Roberts L, Van Remmen H, Epstein CJ, Huang TT. CuZnSOD deficiency leads to persistent and widespread oxidative damage and hepatocarcinogenesis later in life. Oncogene. 2005;24:367–380. - PubMed
-
- Folz RJ, Crapo JD. Extracellular superoxide dismutase (SOD3): tissue-specific expression, genomic characterization, and computer-assisted sequence analysis of the human EC SOD gene. Genomics. 1994;22:162–171. - PubMed
-
- McCord JM, Fridovich I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein) J Biol Chem. 1969;244:6049–6055. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
