

PecS is a global regulator of the symptomatic phase in the phytopathogenic bacterium Erwinia chrysanthemi 3937
- PMID: 18790868
- PMCID: PMC2576657
- DOI: 10.1128/JB.00553-08
PecS is a global regulator of the symptomatic phase in the phytopathogenic bacterium Erwinia chrysanthemi 3937
Abstract
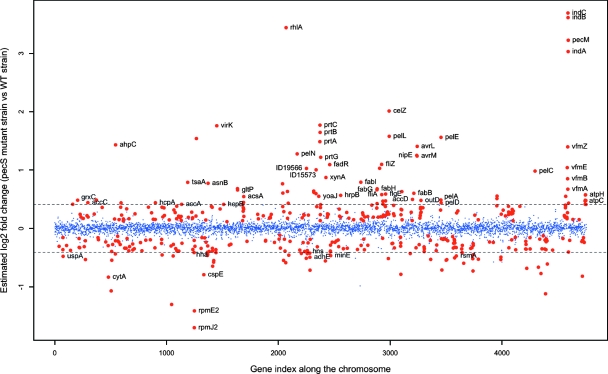
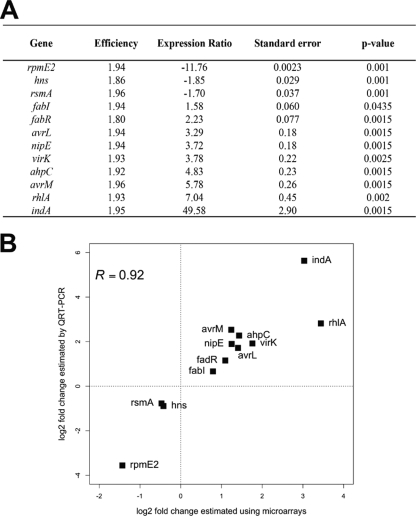
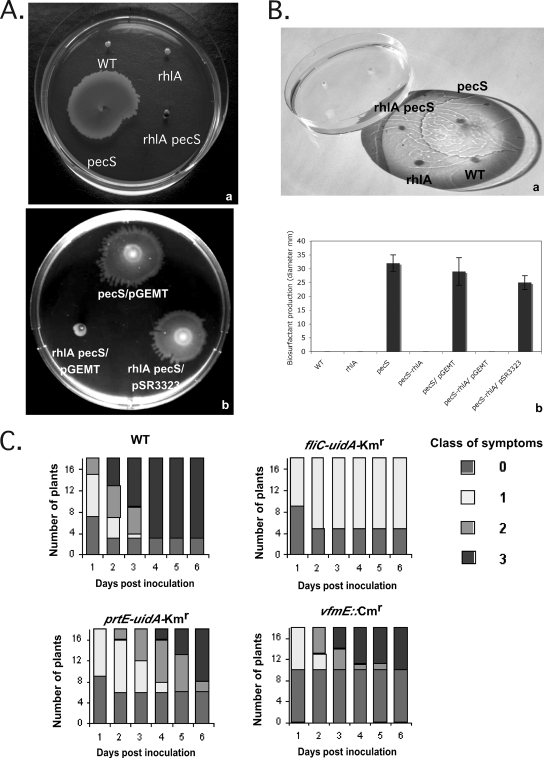
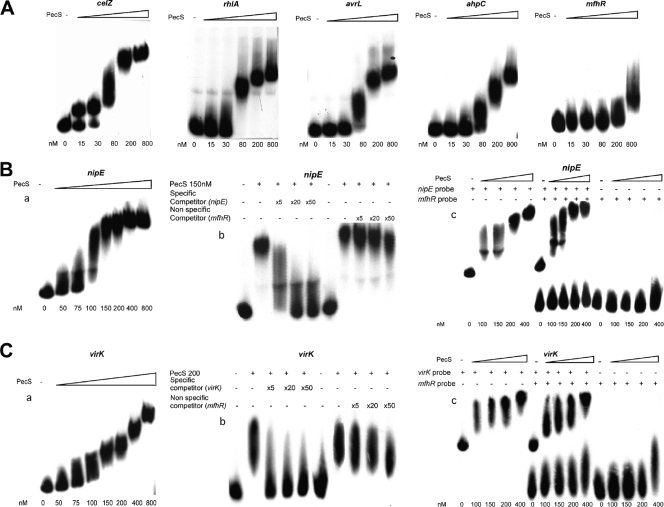
Pathogenicity of the enterobacterium Erwinia chrysanthemi (Dickeya dadantii), the causative agent of soft-rot disease in many plants, is a complex process involving several factors whose production is subject to temporal regulation during infection. PecS is a transcriptional regulator that controls production of various virulence factors. Here, we used microarray analysis to define the PecS regulon and demonstrated that PecS notably regulates a wide range of genes that could be linked to pathogenicity and to a group of genes concerned with evading host defenses. Among the targets are the genes encoding plant cell wall-degrading enzymes and secretion systems and the genes involved in flagellar biosynthesis, biosurfactant production, and the oxidative stress response, as well as genes encoding toxin-like factors such as NipE and hemolysin-coregulated proteins. In vitro experiments demonstrated that PecS interacts with the regulatory regions of five new targets: an oxidative stress response gene (ahpC), a biosurfactant synthesis gene (rhlA), and genes encoding exported proteins related to other plant-associated bacterial proteins (nipE, virK, and avrL). The pecS mutant provokes symptoms more rapidly and with more efficiency than the wild-type strain, indicating that PecS plays a critical role in the switch from the asymptomatic phase to the symptomatic phase. Based on this, we propose that the temporal regulation of the different groups of genes required for the asymptomatic phase and the symptomatic phase is, in part, the result of a gradual modulation of PecS activity triggered during infection in response to changes in environmental conditions emerging from the interaction between both partners.
Figures

References
-
- Andrews, J. 2001. Determination of minimum inhibitory concentrations J. Antimicrob. Chemother. 485-16. - PubMed
-
- Bardonnet, N., and C. Blanco. 1992. ′uidA-antibiotic-resistance cassettes for insertion mutagenesis, gene fusions and genetic constructions. FEMS Microbiol. Lett. 72243-248. - PubMed
-
- Beaulieu, C., M. Boccara, and F. Vangijsegem. 1993. Pathogenic behavior of pectinase-defective Erwinia chrysanthemi mutants on different plants. Mol. Plant-Microbe Interact. 6197-202.
-
- Benjamini, Y., and Y. Hochberg. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. Royal Stat. Soc. Ser. B 57289-300.
-
- Boccara, M., A. Diolez, M. Rouve, and A. Kotoujansky. 1988. The role of the individual pectate lyases of Erwinia chrysanthemi strain 3937 in pathogenicity on Saintpaulia plants. Physiol. Mol. Plant Pathol. 3395-104.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
