

Identification and characterization of cyclic diguanylate signaling systems controlling rugosity in Vibrio cholerae
- PMID: 18790873
- PMCID: PMC2576663
- DOI: 10.1128/JB.00564-08
Identification and characterization of cyclic diguanylate signaling systems controlling rugosity in Vibrio cholerae
Abstract
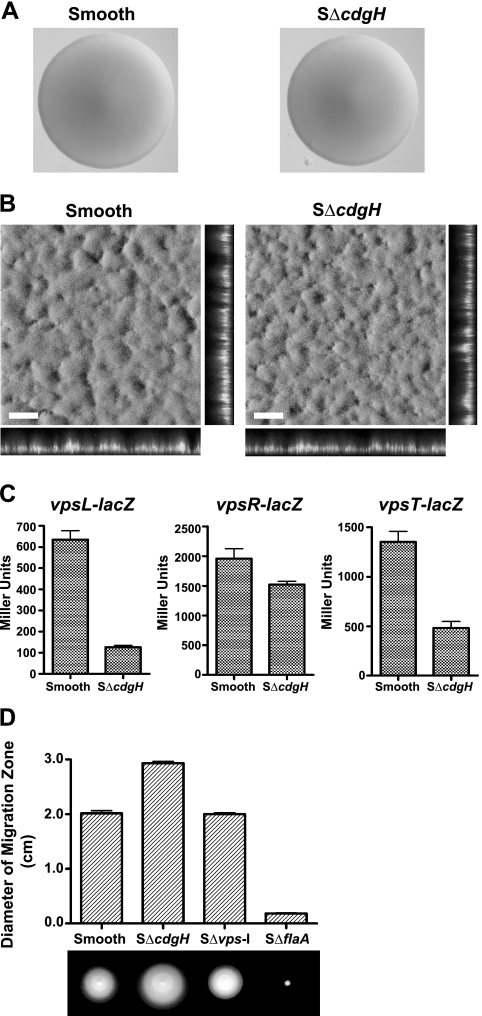
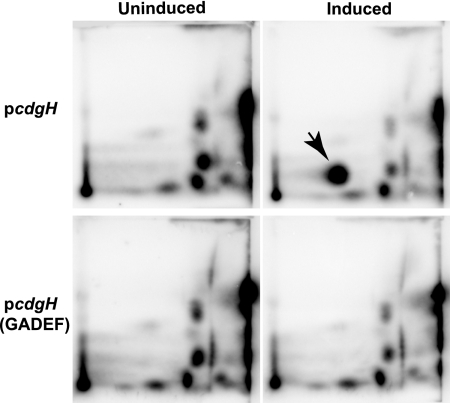
Vibrio cholerae, the causative agent of the disease cholera, can generate rugose variants that have an increased capacity to form biofilms. Rugosity and biofilm formation are critical for the environmental survival and transmission of the pathogen, and these processes are controlled by cyclic diguanylate (c-di-GMP) signaling systems. c-di-GMP is produced by diguanylate cyclases (DGCs) and degraded by phosphodiesterases (PDEs). Proteins that contain GGDEF domains act as DGCs, whereas proteins that contain EAL or HD-GYP domains act as PDEs. In the V. cholerae genome there are 62 genes that are predicted to encode proteins capable of modulating the cellular c-di-GMP concentration. We previously identified two DGCs, VpvC and CdgA, that can control the switch between smooth and rugose. To identify other c-di-GMP signaling proteins involved in rugosity, we generated in-frame deletion mutants of all genes predicted to encode proteins with GGDEF and EAL domains and then searched for mutants with altered rugosity. In this study, we identified two new genes, cdgG and cdgH, involved in rugosity control. We determined that CdgH acts as a DGC and positively regulates rugosity, whereas CdgG does not have DGC activity and negatively regulates rugosity. In addition, epistasis analysis with CdgG, CdgH, and other DGCs and PDEs controlling rugosity revealed that CdgG and CdgH act in parallel with previously identified c-di-GMP signaling proteins to control rugosity in V. cholerae. We also determined that PilZ domain-containing c-di-GMP binding proteins contribute minimally to rugosity, indicating that there are additional c-di-GMP binding proteins controlling rugosity in V. cholerae.
Figures










References
-
- Amikam, D., and M. Y. Galperin. 2006. PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics 223-6. - PubMed
-
- Bao, Y., D. P. Lies, H. Fu, and G. P. Roberts. 1991. An improved Tn7-based system for the single-copy insertion of cloned genes into chromosomes of gram-negative bacteria. Gene 109167-168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
