

The ATG autophagic conjugation system in maize: ATG transcripts and abundance of the ATG8-lipid adduct are regulated by development and nutrient availability
- PMID: 18790996
- PMCID: PMC2613746
- DOI: 10.1104/pp.108.126714
The ATG autophagic conjugation system in maize: ATG transcripts and abundance of the ATG8-lipid adduct are regulated by development and nutrient availability
Abstract
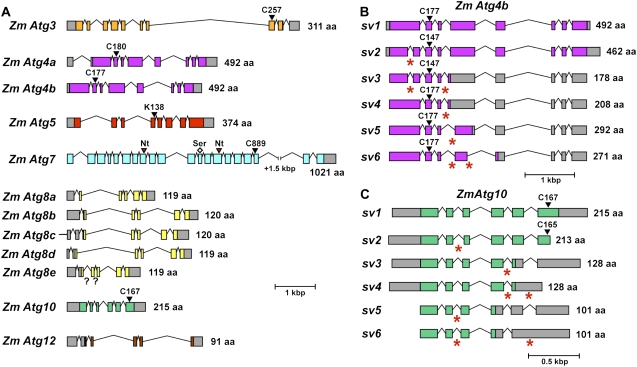
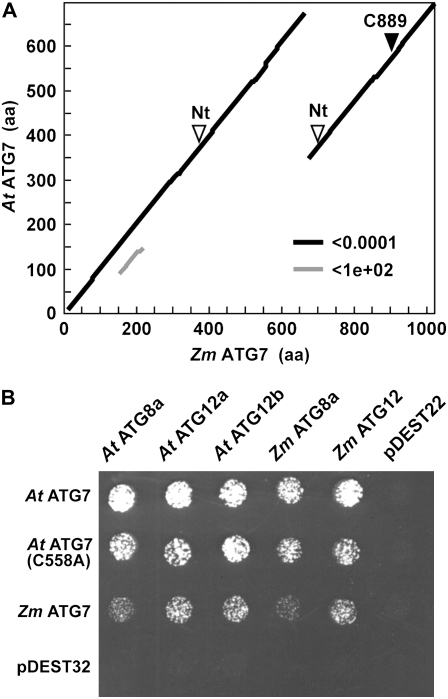
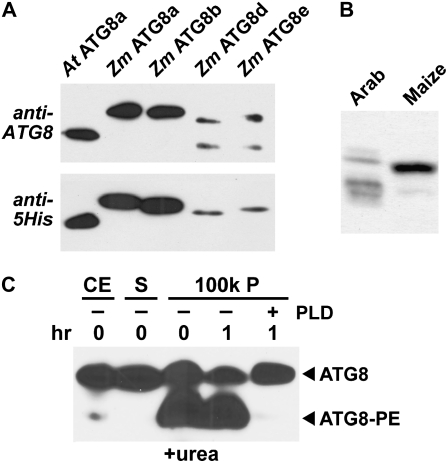
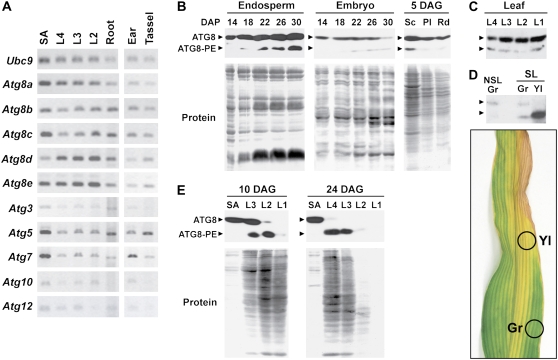
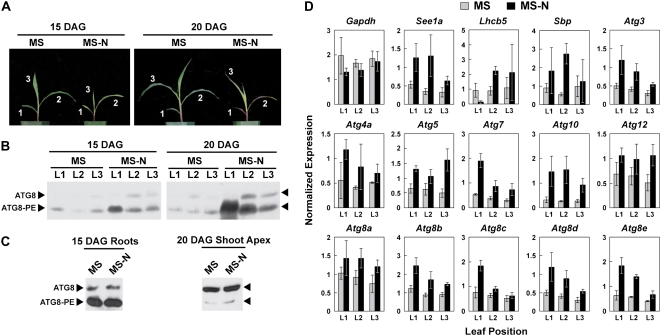
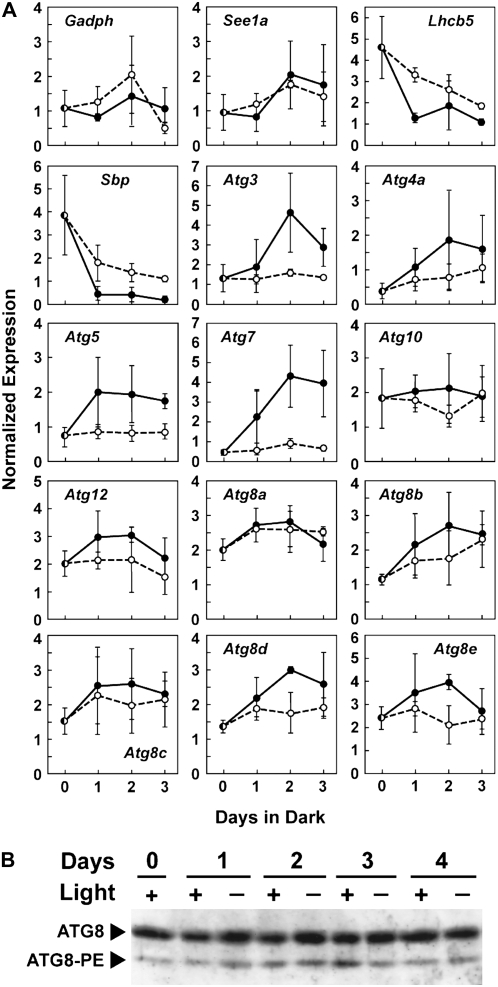
Plants employ sophisticated mechanisms to recycle intracellular constituents needed for growth, development, and survival under nutrient-limiting conditions. Autophagy is one important route in which cytoplasm and organelles are sequestered in bulk into vesicles and subsequently delivered to the vacuole for breakdown by resident hydrolases. The formation and trafficking of autophagic vesicles are directed in part by associated conjugation cascades that couple the AUTOPHAGY-RELATED8 (ATG8) and ATG12 proteins to their respective targets, phosphatidylethanolamine and the ATG5 protein. To help understand the importance of autophagy to nutrient remobilization in cereals, we describe here the ATG8/12 conjugation cascades in maize (Zea mays) and examine their dynamics during development, leaf senescence, and nitrogen and fixed-carbon starvation. From searches of the maize genomic sequence using Arabidopsis (Arabidopsis thaliana) and rice (Oryza sativa) counterparts as queries, we identified orthologous loci encoding all components necessary for ATG8/12 conjugation, including a five-member gene family expressing ATG8. Alternative splicing was evident for almost all Atg transcripts, which could have important regulatory consequences. In addition to free ATG8, its membrane-associated, lipidated form was detected in many maize tissues, suggesting that its conjugation cascade is active throughout the plant at most, if not all, developmental stages. Levels of Atg transcripts and/or the ATG8-phosphatidylethanolamine adduct increase during leaf senescence and nitrogen and fixed-carbon limitations, indicating that autophagy plays a key role in nutrient remobilization. The description of the maize ATG system now provides a battery of molecular and biochemical tools to study autophagy in this crop under field conditions.
Figures


References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215 403–410 - PubMed
-
- Bassham DC (2007) Plant autophagy: more than a starvation response. Curr Opin Plant Biol 10 587–593 - PubMed
-
- Buchanan-Wollaston V, Page T, Harrison E, Breeze E, Lim PO, Nam HG, Lin JF, Wu SH, Swidzinski J, Ishizaki K, et al (2005) Comparative transcriptome analysis reveals significant differences in gene expression and signalling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J 42 567–585 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
