

Glucose deprivation inhibits multiple key gene expression events and effector functions in CD8+ T cells
- PMID: 18792400
- PMCID: PMC3008428
- DOI: 10.1002/eji.200838289
Glucose deprivation inhibits multiple key gene expression events and effector functions in CD8+ T cells
Abstract
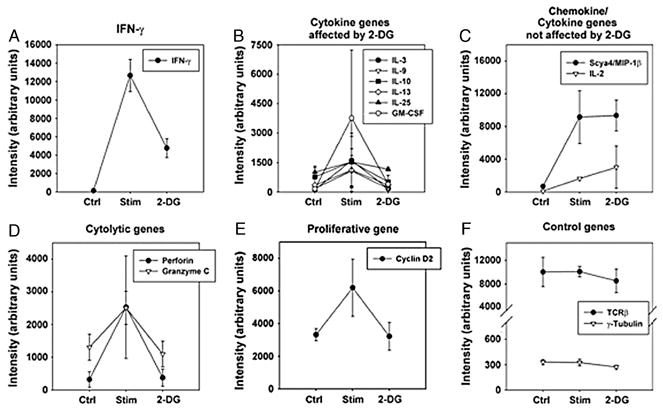
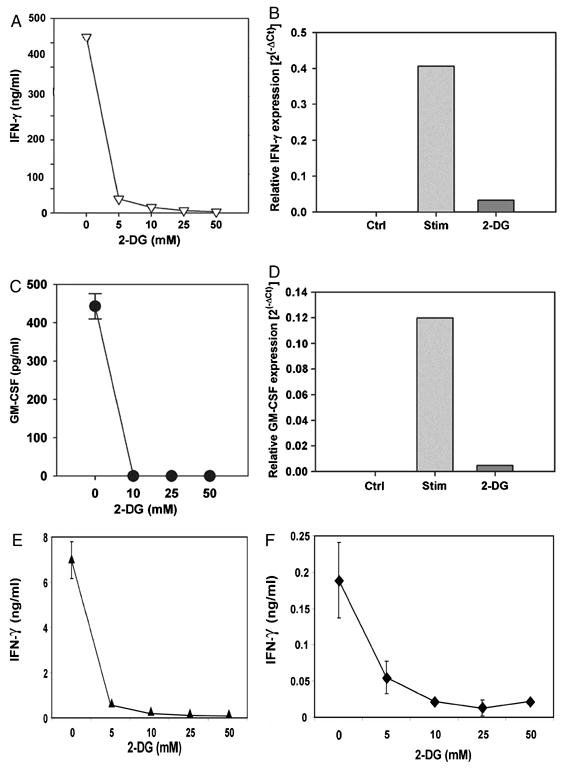
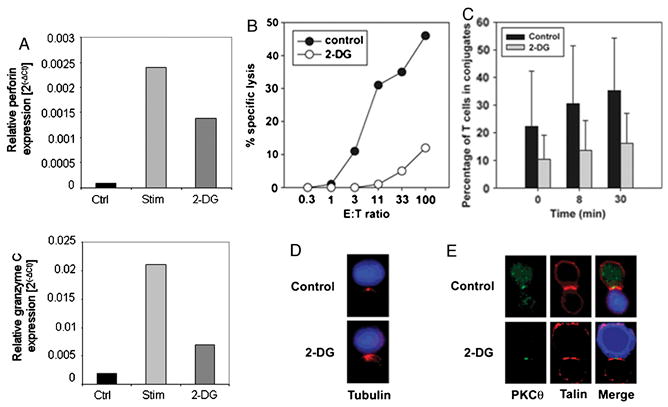
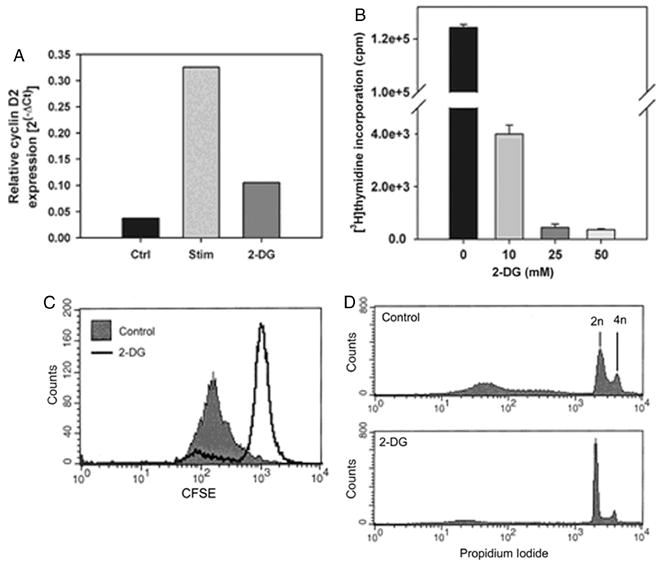
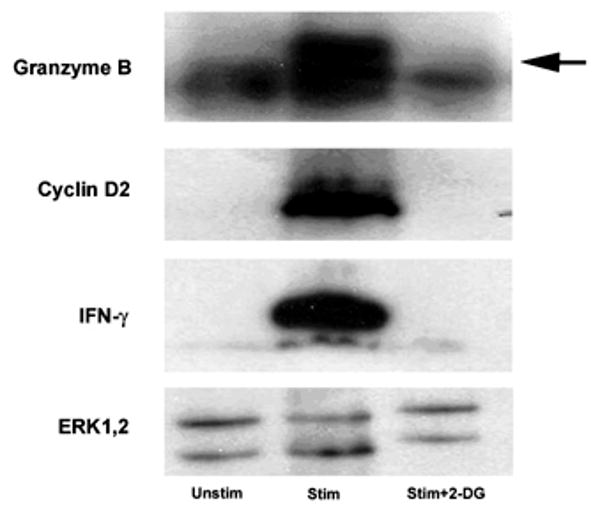
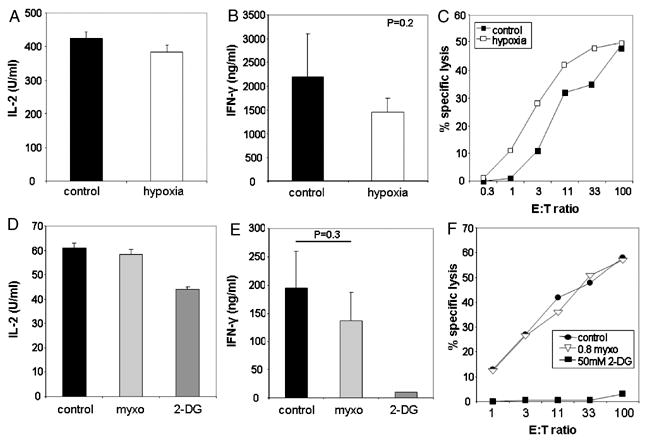
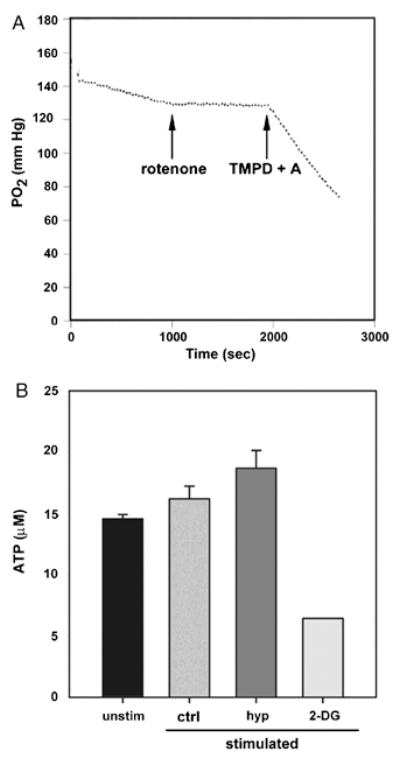
We recently reported that differentiation of CD8(+) T cells from the naïve to the effector state involves the upregulation of glucose-dependent metabolism. Glucose deprivation or inhibition of glycolysis by 2-deoxy-D-glucose (2-DG) selectively inhibited production of IFN-gamma but not of IL-2. To determine a more global role of glucose metabolism on effector T-cell function, we performed gene array analysis on CD8(+) effector T cells stimulated in the presence or absence of 2-DG. We observed that expression of only 10% of genes induced by TCR/CD28 signaling was inhibited by 2-DG. Among these were genes for key cytokines, cell cycle molecules, and cytotoxic granule proteins. Consistent with these results, production of IFN-gamma and GM-CSF, cell cycle progression, upregulation of cyclin D2 protein, cytolytic activity, and upregulation of granzyme B protein and also conjugate formation were exquisitely glucose-dependent. In contrast to glucose, oxygen was little utilized by CD8(+) effector T cells, and relative oxygen deprivation did not inhibit these CTL functional properties. Our results indicate a particularly critical role for glucose in regulating specific effector functions of CD8(+) T cells and have implications for the maintenance of the effector phase of cellular immune responses in target tissue microenvironments such as a solid tumor.
Conflict of interest statement
Figures
References
-
- Cham CM, Gajewski TF. Glucose availability regulates IFN-gamma production and p70S6 kinase activation in CD8+ effector T cells. J Immunol. 2005;174:4670–4677. - PubMed
-
- Fox CJ, Hammerman PS, Thompson CB. Fuel feeds function: energy metabolism and the T-cell response. Nat Rev Immunol. 2005;5:844–852. - PubMed
-
- Stinchcombe JC, Bossi G, Booth S, Griffiths GM. The immunological synapse of CTL contains a secretory domain and membrane bridges. Immunity. 2001;15:751–761. - PubMed
-
- Monks CR, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 1998;395:82–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
