

Effector caspase Dcp-1 and IAP protein Bruce regulate starvation-induced autophagy during Drosophila melanogaster oogenesis
- PMID: 18794330
- PMCID: PMC2542474
- DOI: 10.1083/jcb.200712091
Effector caspase Dcp-1 and IAP protein Bruce regulate starvation-induced autophagy during Drosophila melanogaster oogenesis
Abstract
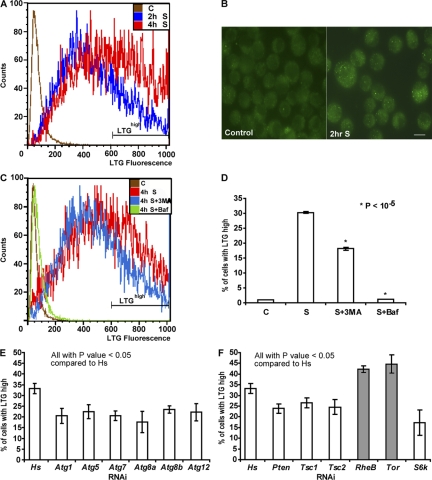
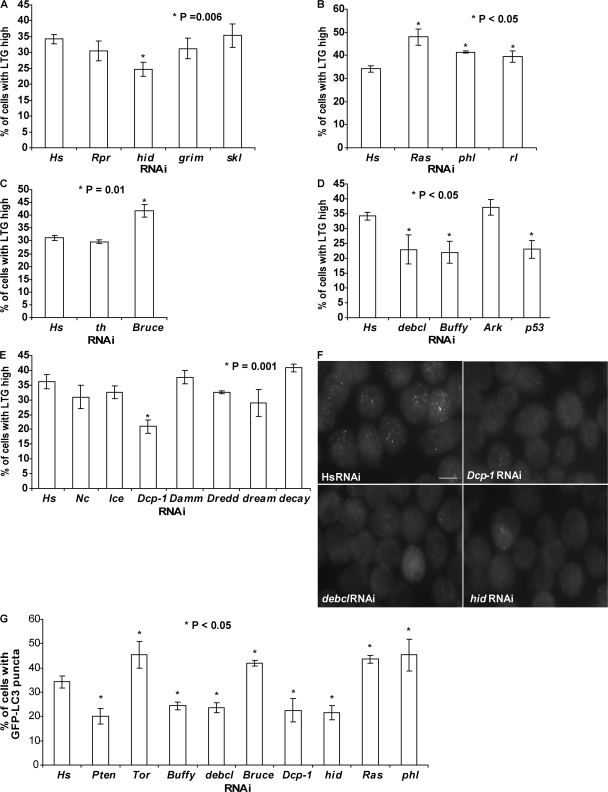
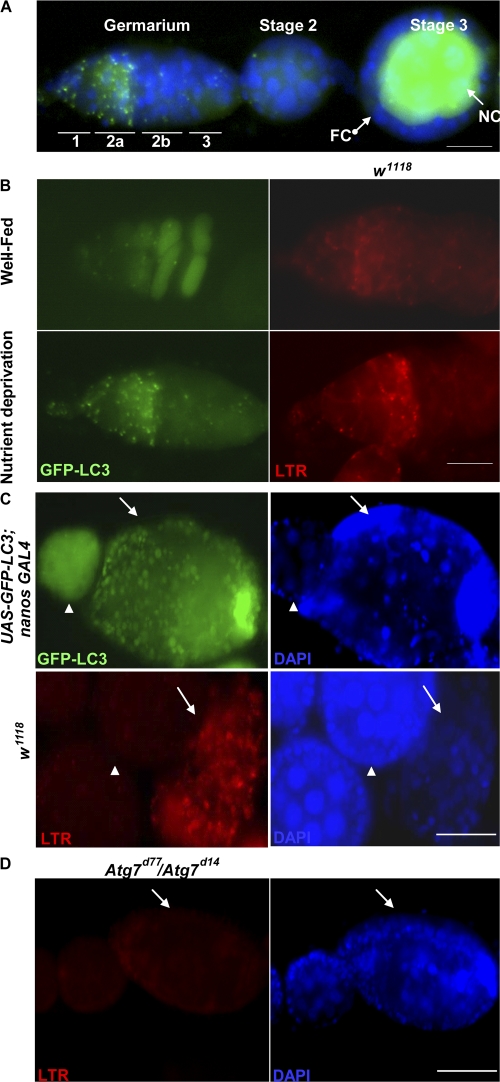
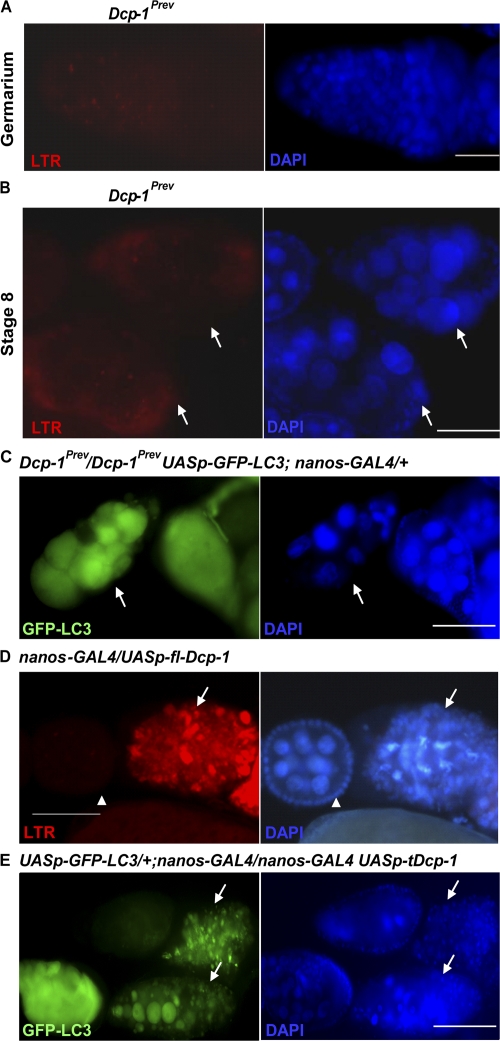
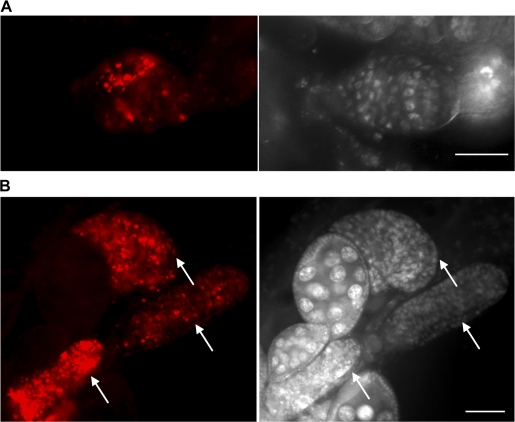
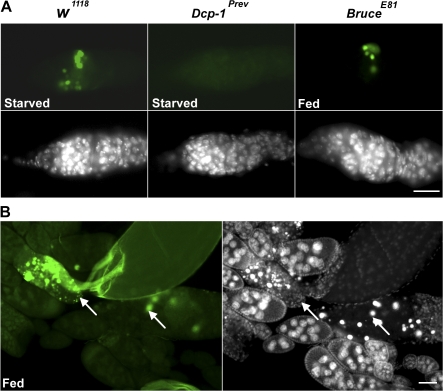
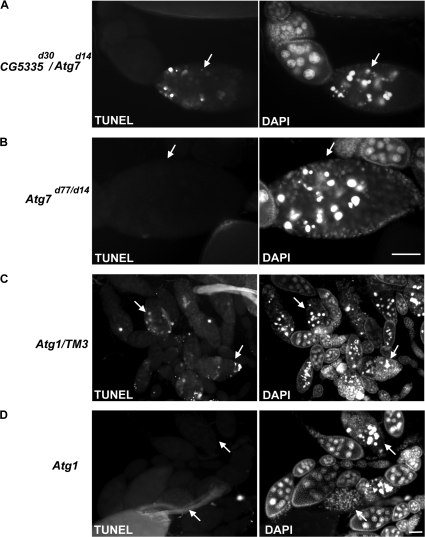
A complex relationship exists between autophagy and apoptosis, but the regulatory mechanisms underlying their interactions are largely unknown. We conducted a systematic study of Drosophila melanogaster cell death-related genes to determine their requirement in the regulation of starvation-induced autophagy. We discovered that six cell death genes--death caspase-1 (Dcp-1), hid, Bruce, Buffy, debcl, and p53-as well as Ras-Raf-mitogen activated protein kinase signaling pathway components had a role in autophagy regulation in D. melanogaster cultured cells. During D. melanogaster oogenesis, we found that autophagy is induced at two nutrient status checkpoints: germarium and mid-oogenesis. At these two stages, the effector caspase Dcp-1 and the inhibitor of apoptosis protein Bruce function to regulate both autophagy and starvation-induced cell death. Mutations in Atg1 and Atg7 resulted in reduced DNA fragmentation in degenerating midstage egg chambers but did not appear to affect nuclear condensation, which indicates that autophagy contributes in part to cell death in the ovary. Our study provides new insights into the molecular mechanisms that coordinately regulate autophagic and apoptotic events in vivo.
Figures
References
-
- Arama, E., J. Agapite, and H. Steller. 2003. Caspase activity and a specific cytochrome C are required for sperm differentiation in Drosophila. Dev. Cell. 4:687–697. - PubMed
-
- Baum, J.S., E. Arama, H. Steller, and K. McCall. 2007. The Drosophila caspases Strica and Dronc function redundantly in programmed cell death during oogenesis. Cell Death Differ. 14:1508–1517. - PubMed
-
- Bergmann, A., J. Agapite, K. McCall, and H. Steller. 1998. The Drosophila gene hid is a direct molecular target of Ras-dependent survival signaling. Cell. 95:331–341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
