

Defining the regulatory network of the tissue-specific splicing factors Fox-1 and Fox-2
- PMID: 18794351
- PMCID: PMC2546699
- DOI: 10.1101/gad.1703108
Defining the regulatory network of the tissue-specific splicing factors Fox-1 and Fox-2
Erratum in
- Genes Dev. 2008 Oct 15;22(20):2902
Abstract
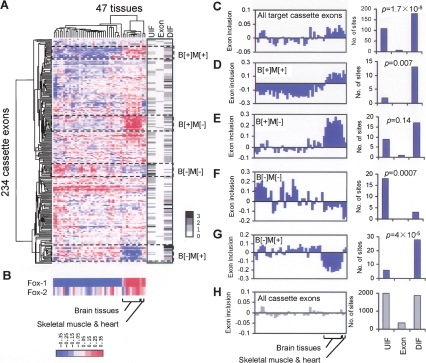
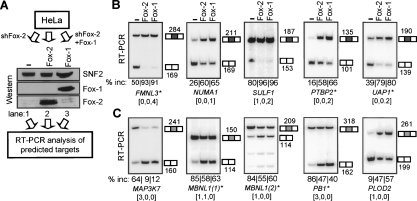
The precise regulation of many alternative splicing (AS) events by specific splicing factors is essential to determine tissue types and developmental stages. However, the molecular basis of tissue-specific AS regulation and the properties of splicing regulatory networks (SRNs) are poorly understood. Here we comprehensively predict the targets of the brain- and muscle-specific splicing factor Fox-1 (A2BP1) and its paralog Fox-2 (RBM9) and systematically define the corresponding SRNs genome-wide. Fox-1/2 are conserved from worm to human, and specifically recognize the RNA element UGCAUG. We integrate Fox-1/2-binding specificity with phylogenetic conservation, splicing microarray data, and additional computational and experimental characterization. We predict thousands of Fox-1/2 targets with conserved binding sites, at a false discovery rate (FDR) of approximately 24%, including many validated experimentally, suggesting a surprisingly extensive SRN. The preferred position of the binding sites differs according to AS pattern, and determines either activation or repression of exon recognition by Fox-1/2. Many predicted targets are important for neuromuscular functions, and have been implicated in several genetic diseases. We also identified instances of binding site creation or loss in different vertebrate lineages and human populations, which likely reflect fine-tuning of gene expression regulation during evolution.
Figures




References
-
- Bhalla K., Phillips H.A., Crawford J., McKenzie O.L.D., Mulley J.C., Eyre H., Gardner A.E., Kremmidiotis G., Callen D.F. The de novo chromosome 16 translocations of two patients with abnormal phenotypes (mental retardation and epilepsy) disrupt the A2BP1 gene. J. Hum. Genet. 2004;49:308–311. - PubMed
-
- Black D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003;72:291–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials