

Role of the N- and C-lobes of calmodulin in the activation of Ca(2+)/calmodulin-dependent protein kinase II
- PMID: 18795794
- PMCID: PMC2665295
- DOI: 10.1021/bi8007033
Role of the N- and C-lobes of calmodulin in the activation of Ca(2+)/calmodulin-dependent protein kinase II
Abstract
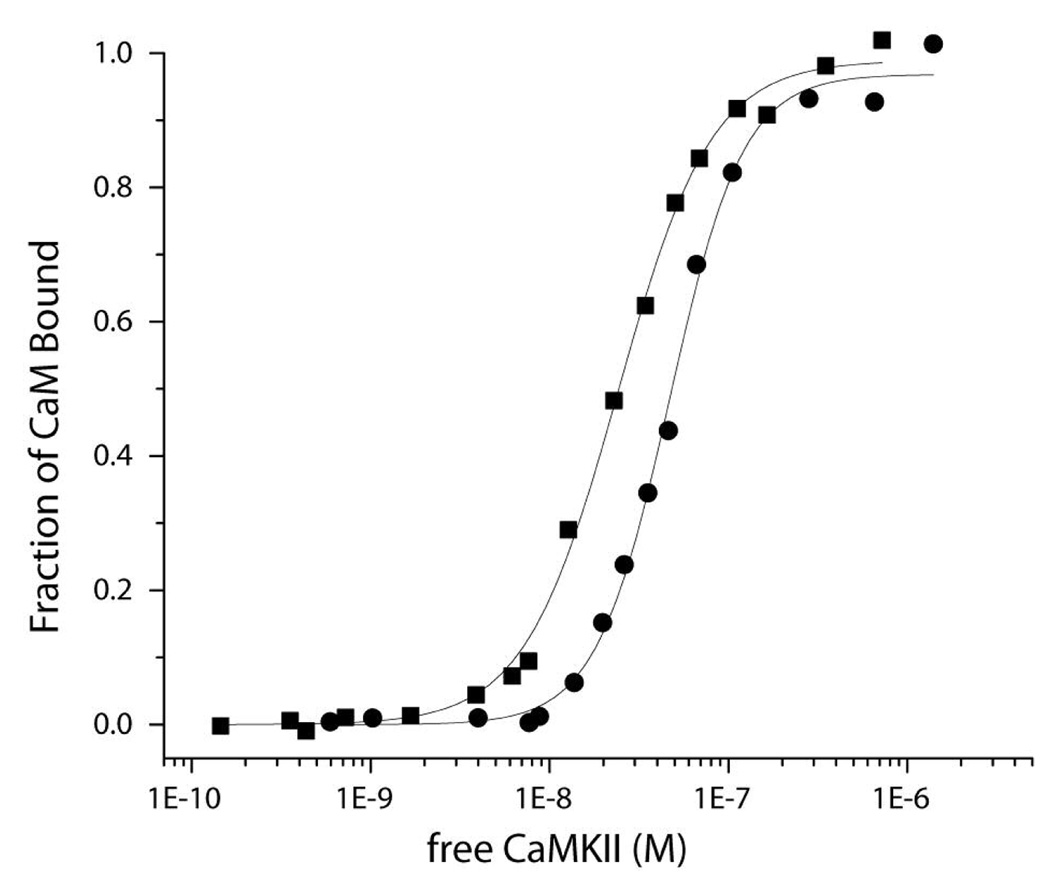
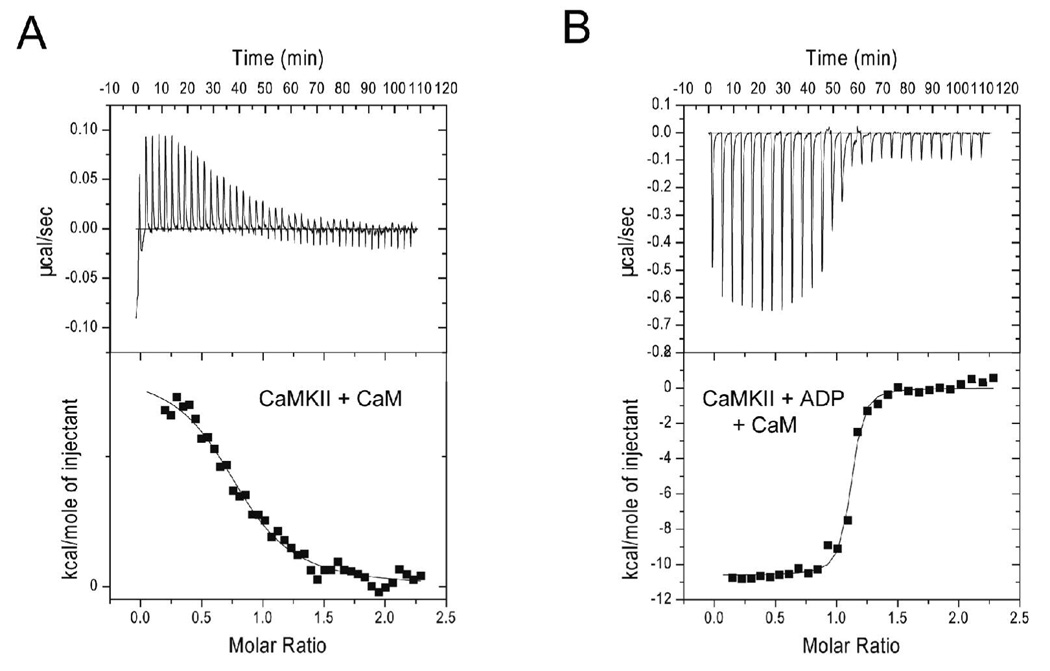
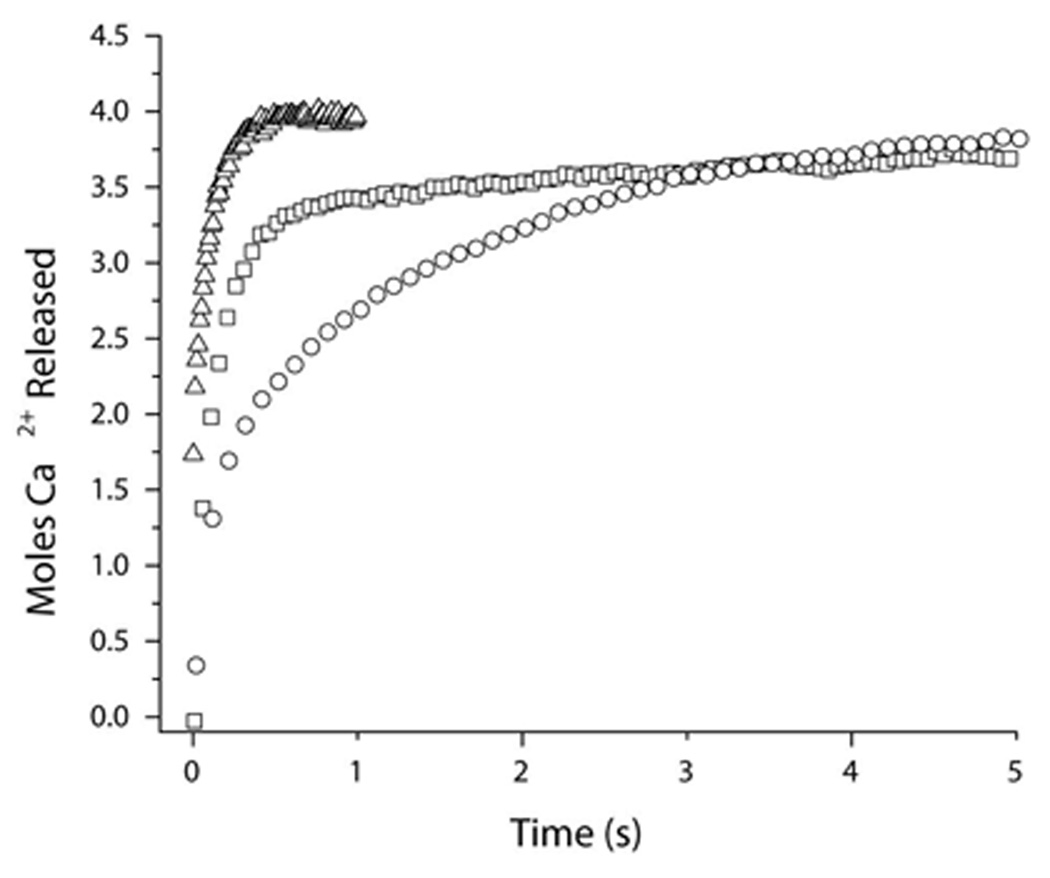
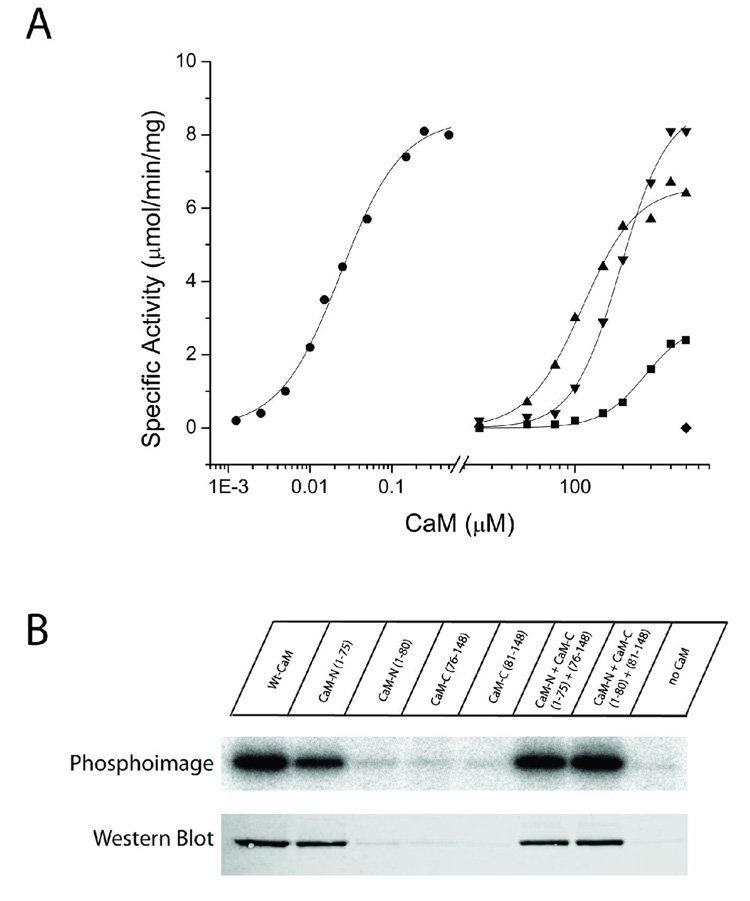
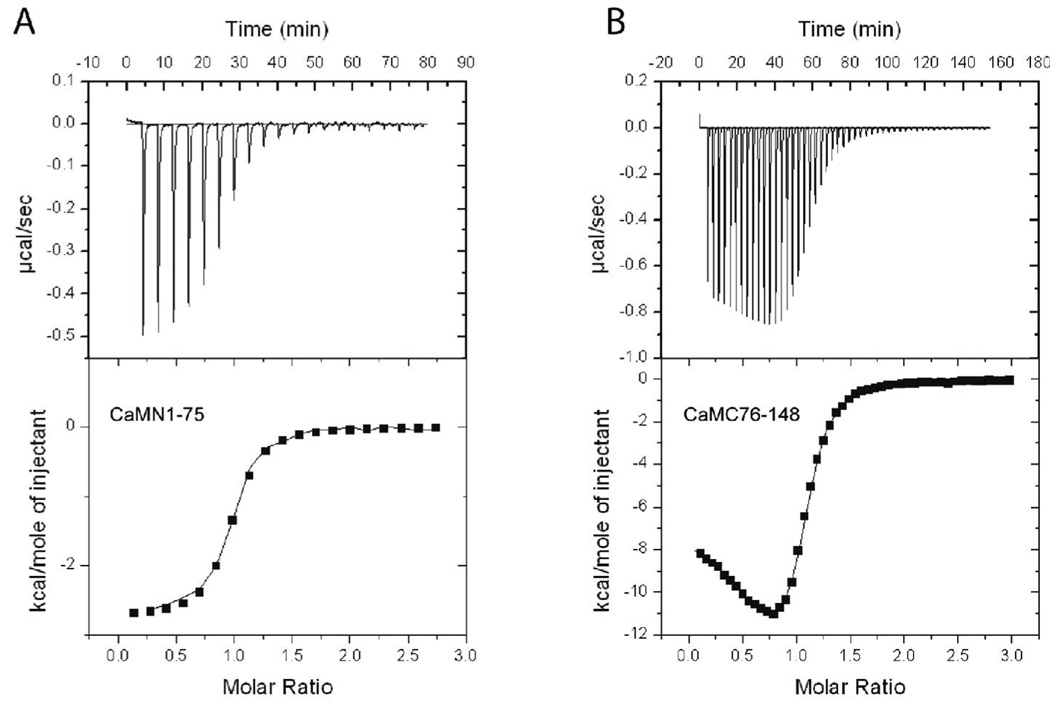
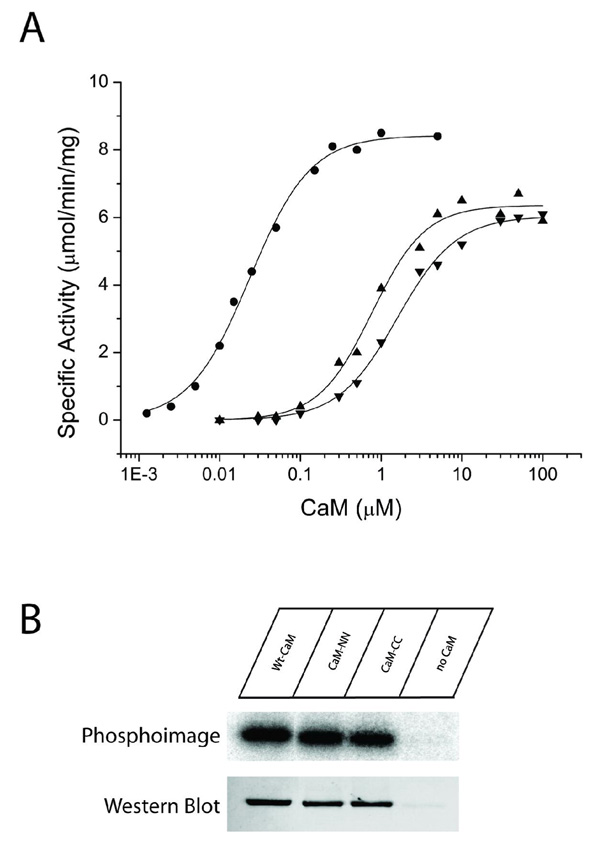
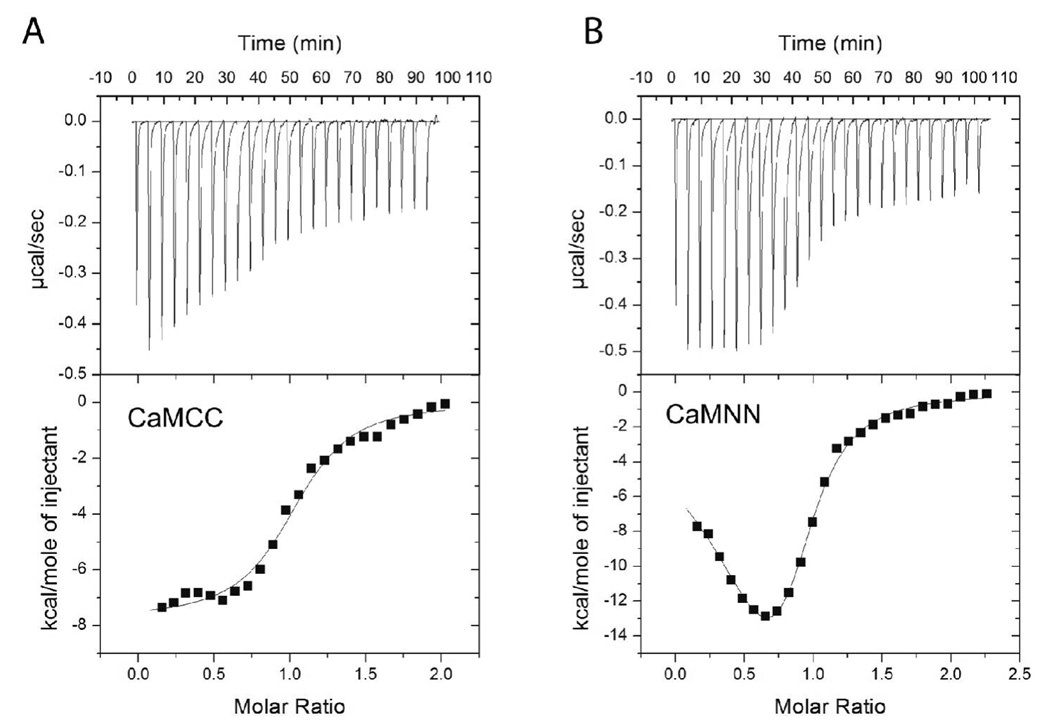
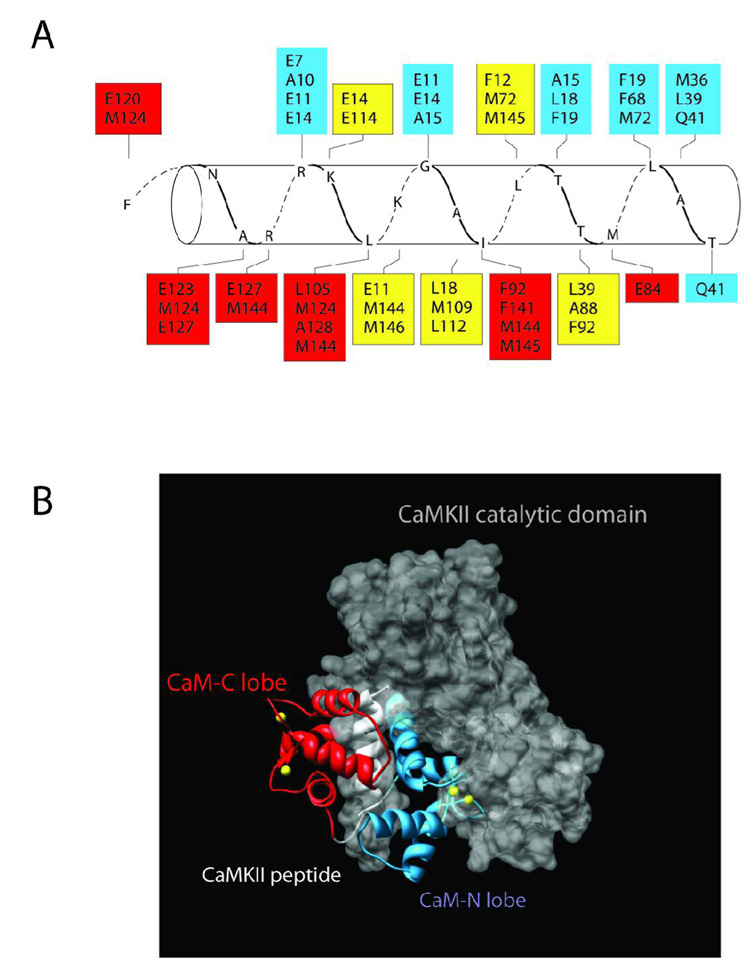
Understanding the principles of calmodulin (CaM) activation of target enzymes will help delineate how this seemingly simple molecule can play such a complex role in transducing Ca (2+)-signals to a variety of downstream pathways. In the work reported here, we use biochemical and biophysical tools and a panel of CaM constructs to examine the lobe specific interactions between CaM and CaMKII necessary for the activation and autophosphorylation of the enzyme. Interestingly, the N-terminal lobe of CaM by itself was able to partially activate and allow autophosphorylation of CaMKII while the C-terminal lobe was inactive. When used together, CaMN and CaMC produced maximal CaMKII activation and autophosphorylation. Moreover, CaMNN and CaMCC (chimeras of the two N- or C-terminal lobes) both activated the kinase but with greater K act than for wtCaM. Isothermal titration calorimetry experiments showed the same rank order of affinities of wtCaM > CaMNN > CaMCC as those determined in the activity assay and that the CaM to CaMKII subunit binding ratio was 1:1. Together, our results lead to a proposed sequential mechanism to describe the activation pathway of CaMKII led by binding of the N-lobe followed by the C-lobe. This mechanism contrasts the typical sequential binding mode of CaM with other CaM-dependent enzymes, where the C-lobe of CaM binds first. The consequence of such lobe specific binding mechanisms is discussed in relation to the differential rates of Ca (2+)-binding to each lobe of CaM during intracellular Ca (2+) oscillations.
Figures
References
-
- Hudmon A, Schulman H. Neuronal CA2+/calmodulin-dependent protein kinase II: the role of structure and autoregulation in cellular function. Annu Rev Biochem. 2002;71:473–510. - PubMed
-
- Kolodziej SJ, Hudmon A, Waxham MN, Stoops JK. Three-dimensional reconstructions of calcium/calmodulin-dependent (CaM) kinase IIalpha and truncated CaM kinase IIalpha reveal a unique organization for its structural core and functional domains. J Biol Chem. 2000;275:14354–14359. - PubMed
-
- Miller SG, Kennedy MB. Regulation of brain type II Ca2+/calmodulin-dependent protein kinase by autophosphorylation: a Ca2+-triggered molecular switch. Cell. 1986;44:861–870. - PubMed
-
- Meyer T, Hanson PI, Stryer L, Schulman H. Calmodulin trapping by calcium-calmodulin-dependent protein kinase. Science. 1992;256:1199–1202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
