

A NMDA receptor glycine site partial agonist, GLYX-13, simultaneously enhances LTP and reduces LTD at Schaffer collateral-CA1 synapses in hippocampus
- PMID: 18796308
- PMCID: PMC2661239
- DOI: 10.1016/j.neuropharm.2008.08.018
A NMDA receptor glycine site partial agonist, GLYX-13, simultaneously enhances LTP and reduces LTD at Schaffer collateral-CA1 synapses in hippocampus
Abstract
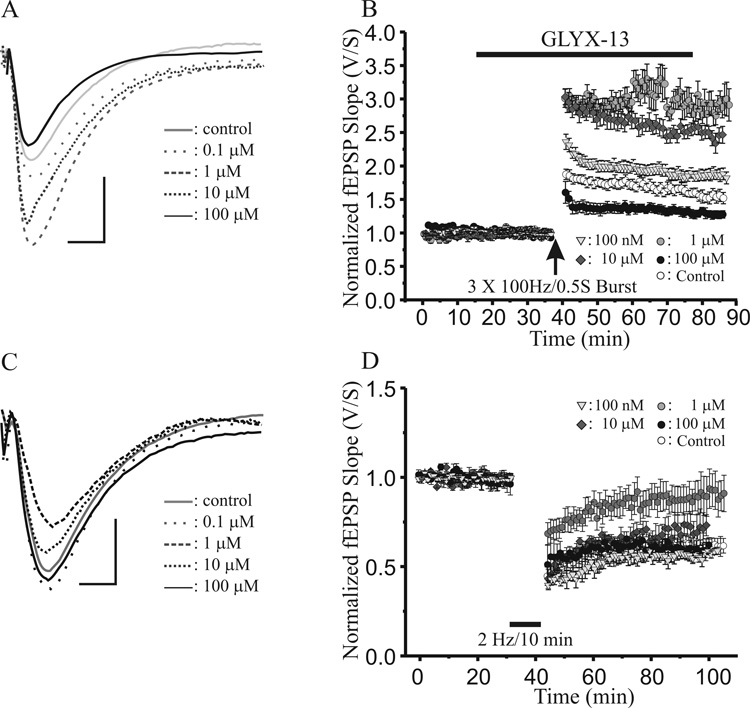
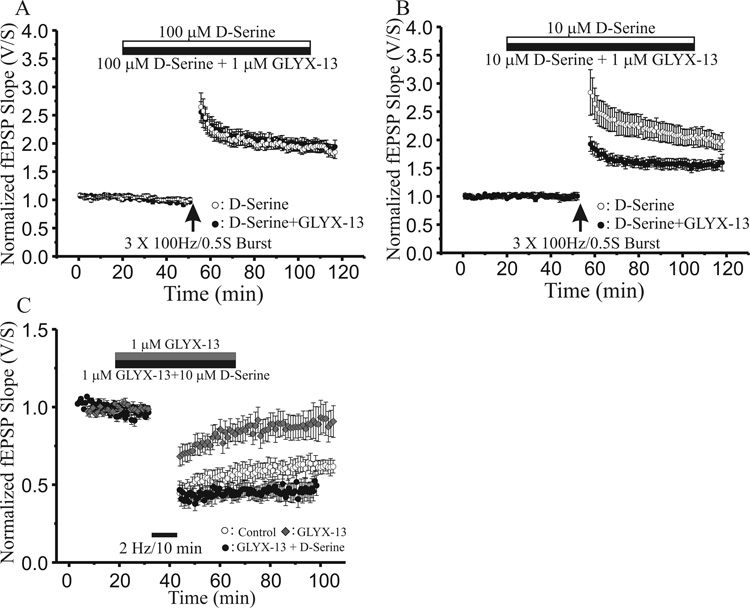
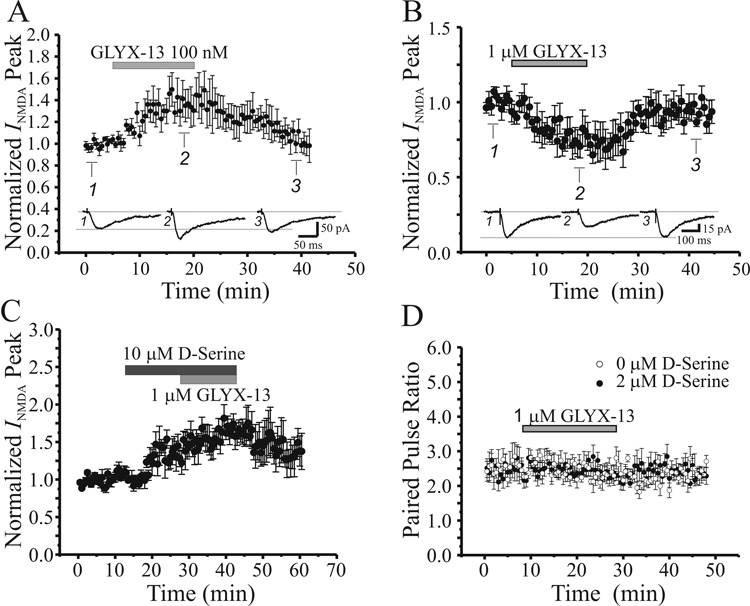
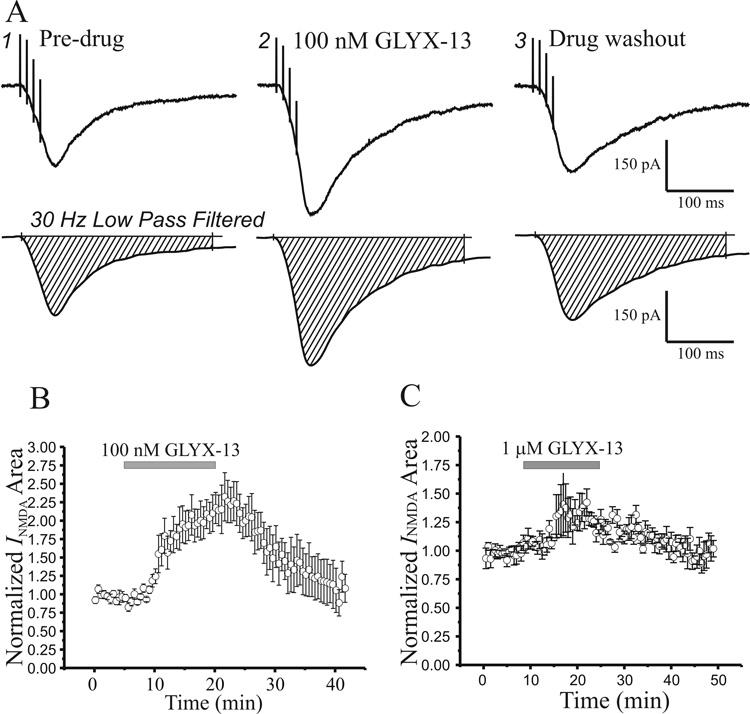
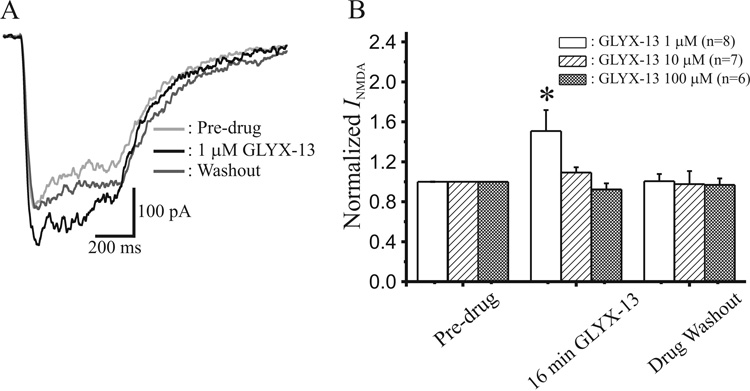
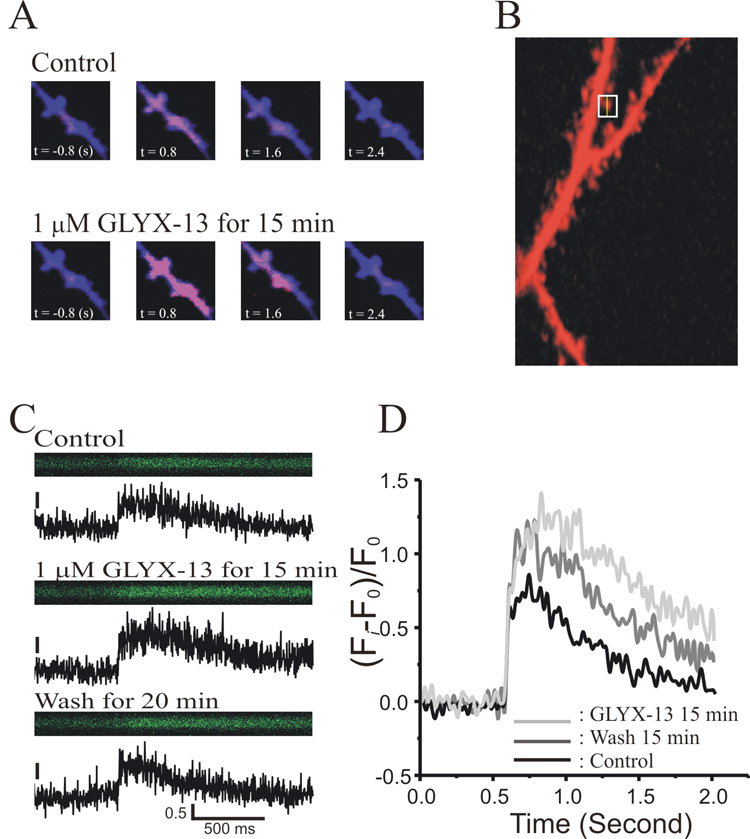
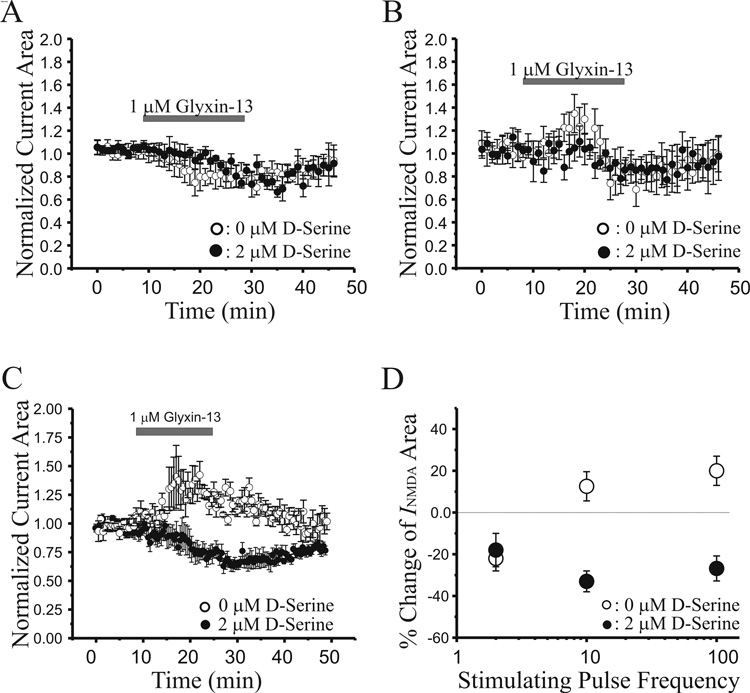
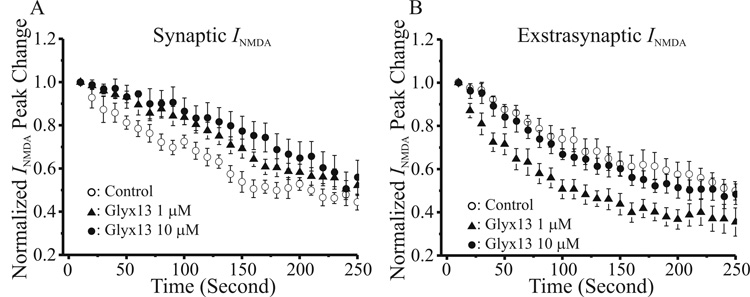
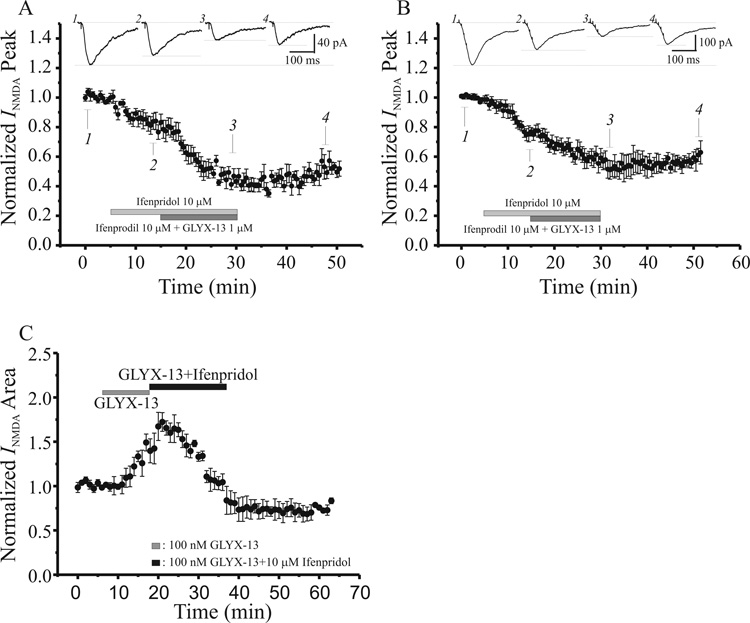
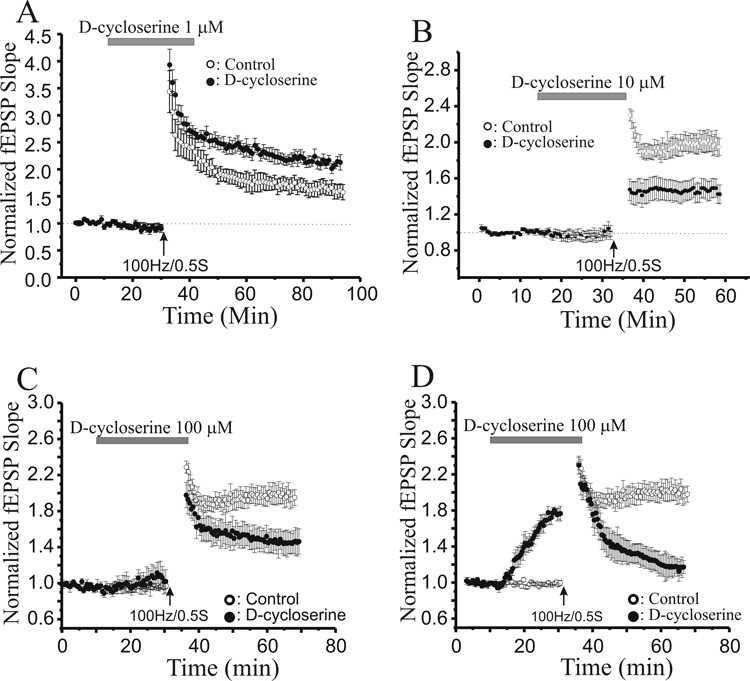
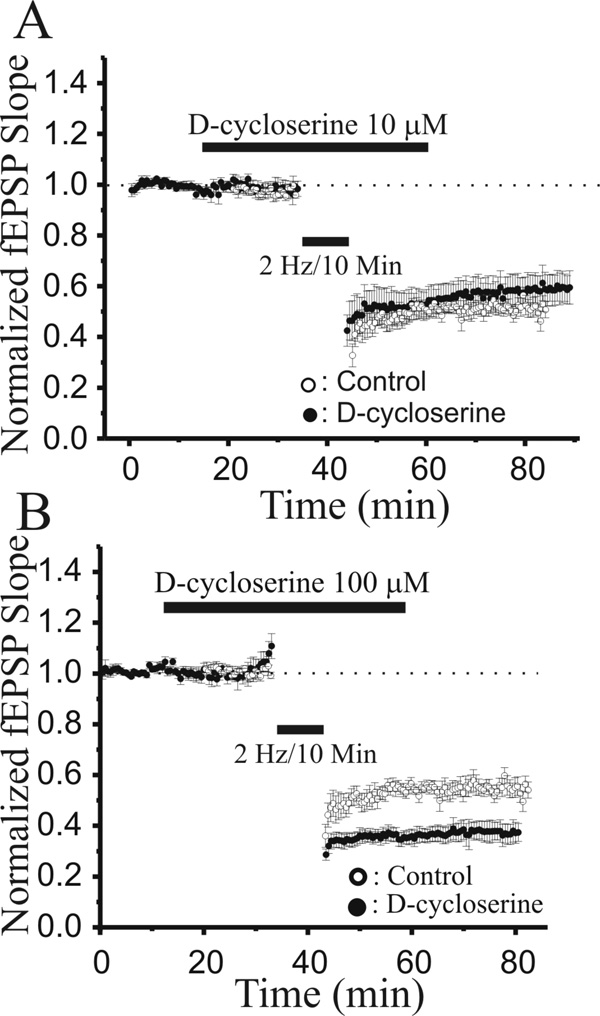
N-methyl-D-aspartate glutamate receptors (NMDARs) are a key route for Ca2+ influx into neurons important to both activity-dependent synaptic plasticity and, when uncontrolled, triggering events that cause neuronal degeneration and death. Among regulatory binding sites on the NMDAR complex is a glycine binding site, distinct from the glutamate binding site, which must be co-activated for NMDAR channel opening. We developed a novel glycine site partial agonist, GLYX-13, which is both nootropic and neuroprotective in vivo. Here, we assessed the effects of GLYX-13 on long-term synaptic plasticity and NMDAR transmission at Schaffer collateral-CA1 synapses in hippocampal slices in vitro. GLYX-13 simultaneously enhanced the magnitude of long-term potentiation (LTP) of synaptic transmission, while reducing long-term depression (LTD). GLYX-13 reduced NMDA receptor-mediated synaptic currents in CA1 pyramidal neurons evoked by low frequency Schaffer collateral stimulation, but enhanced NMDAR currents during high frequency bursts of activity, and these actions were occluded by a saturating concentration of the glycine site agonist d-serine. Direct two-photon imaging of Schaffer collateral burst-evoked increases in [Ca2+] in individual dendritic spines revealed that GLYX-13 selectively enhanced burst-induced NMDAR-dependent spine Ca2+ influx. Examining the rate of MK-801 block of synaptic versus extrasynaptic NMDAR-gated channels revealed that GLYX-13 selectively enhanced activation of burst-driven extrasynaptic NMDARs, with an action that was blocked by the NR2B-selective NMDAR antagonist ifenprodil. Our data suggest that GLYX-13 may have unique therapeutic potential as a learning and memory enhancer because of its ability to simultaneously enhance LTP and suppress LTD.
Figures
References
-
- Akhondzadeh S. Hippocampal synaptic plasticity and cognition. J. Clin. Pharm. Ther. 1999;24:241–248. - PubMed
-
- Asztely F, Erdemli G, Kullmann DM. Extrasynaptic glutamate spillover in the hippocampus: dependence on temperature and the role of active glutamate uptake. Neuron. 1997;18:281–293. - PubMed
-
- Billard JM, Rouaud E. Deficit of NMDA receptor activation in CA1 hippocampal area of aged rats is rescued by D-cycloserine. Eur. J. Neurosci. 2007;25:2260–2268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
