

Several deep-sea mussels and their associated symbionts are able to live both on wood and on whale falls
- PMID: 18796394
- PMCID: PMC2614261
- DOI: 10.1098/rspb.2008.1101
Several deep-sea mussels and their associated symbionts are able to live both on wood and on whale falls
Abstract
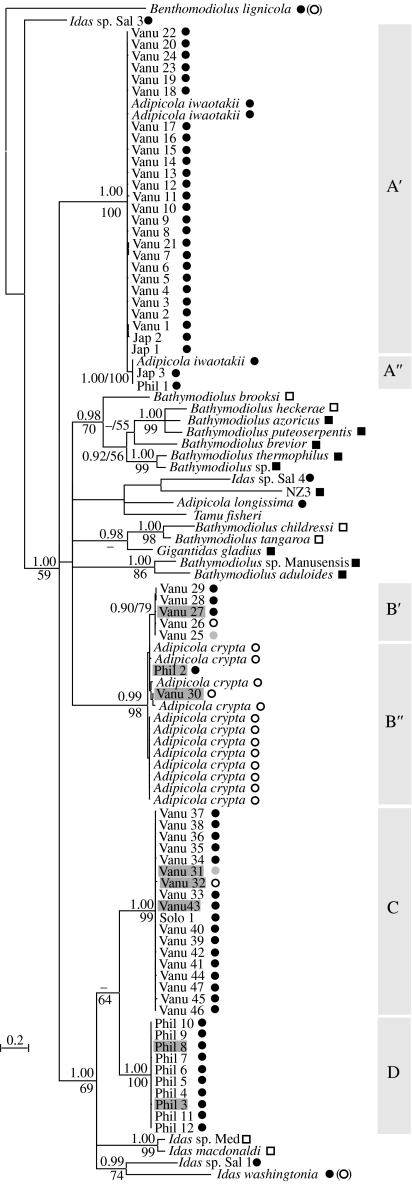
Bathymodiolin mussels occur at hydrothermal vents and cold seeps, where they thrive thanks to symbiotic associations with chemotrophic bacteria. Closely related genera Idas and Adipicola are associated with organic falls, ecosystems that have been suggested as potential evolutionary 'stepping stones' in the colonization of deeper and more sulphide-rich environments. Such a scenario should result from specializations to given environments from species with larger ecological niches. This study provides molecular-based evidence for the existence of two mussel species found both on sunken wood and bones. Each species specifically harbours one bacterial phylotype corresponding to thioautotrophic bacteria related to other bathymodiolin symbionts. Phylogenetic patterns between hosts and symbionts are partially congruent. However, active endocytosis and occurrences of minor symbiont lineages within species which are not their usual host suggest an environmental or horizontal rather than strictly vertical transmission of symbionts. Although the bacteria are close relatives, their localization is intracellular in one mussel species and extracellular in the other, suggesting that habitat choice is independent of the symbiont localization. The variation of bacterial densities in host tissues is related to the substrate on which specimens were sampled and could explain the abilities of host species to adapt to various substrates.
Figures



References
-
- Baco, A. R. 2002 Food-web structure, succession, and phylogenetics on deep-sea whale skeletons. In Oceanography, p. 275. Manoa, HI: University of Hawaii.
-
- Cavanaugh, C., McKiness, Z., Newton, I. & Stewart, F. 2006 Marine chemosynthetic symbioses. In The prokaryotes, pp. 475–507. New York, NY: Springer. (doi:10.1007/0-387-30741-9_18) - DOI
-
- Craddock C, Hoeh W.R, Gustafson R.G, Lutz R.A, Hashimoto J, Vrijenhoek R.J. Evolutionary relationships among deep-sea mytilids (Bivalvia: Mytilidae) from hydrothermal vents and cold-water methane/sulfide seeps. Mar. Biol. 1995;121:477–485. doi:10.1007/BF00349456 - DOI
-
- Dell R.K. Mollusca of the family Mytilidae (Bivalvia) associated with organic remains from deep water off New Zealand, with revisions of the genera Adipicola Dautzenberg, 1927 and Idasola Iredale, 1915. Natl Museum NZ Rec. 1987;3:17–36.
-
- Deming J.W, Reysenbach A.-L, Macko S.A, Smith C.R. Evidence for the microbial basis of a chemoautotrophic invertebrate community at a whale fall on the deep seafloor: bone-colonizing bacteria and invertebrate endosymbionts. Microsc. Res. Tech. 1997;37:162–170. doi:10.1002/(SICI)1097-0029(19970415)37:2<162::AID-JEMT4>3.0.CO;2-Q - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
