

Differential roles of the COOH termini of AAA subunits of PA700 (19 S regulator) in asymmetric assembly and activation of the 26 S proteasome
- PMID: 18796432
- PMCID: PMC2581596
- DOI: 10.1074/jbc.M805935200
Differential roles of the COOH termini of AAA subunits of PA700 (19 S regulator) in asymmetric assembly and activation of the 26 S proteasome
Abstract
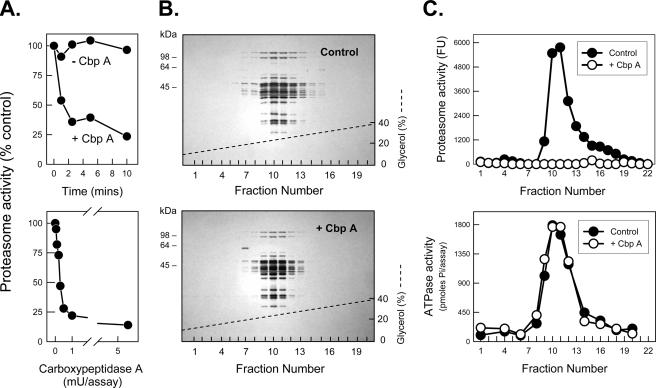
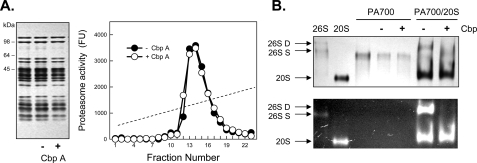
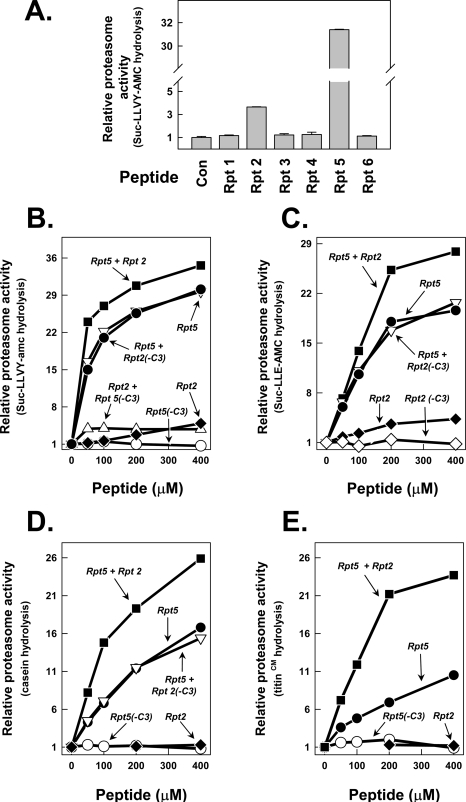
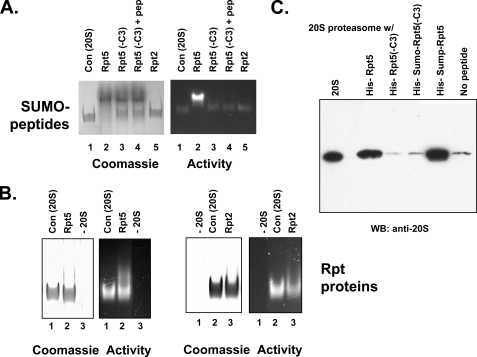
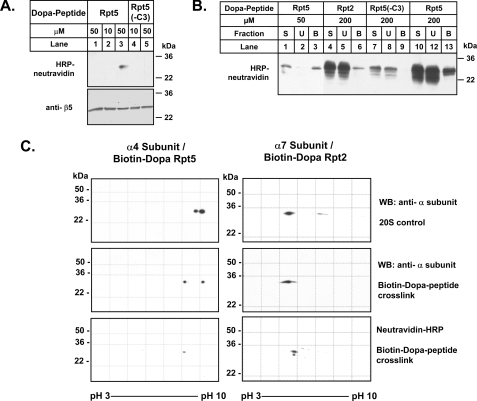
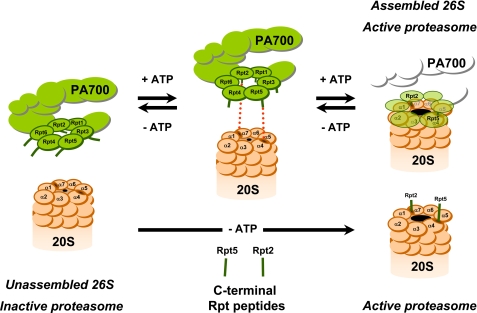
The 26 S proteasome is an energy-dependent protease that degrades proteins modified with polyubiquitin chains. It is assembled from two multi-protein subcomplexes: a protease (20 S proteasome) and an ATPase regulatory complex (PA700 or 19 S regulatory particle) that contains six different AAA family subunits (Rpt1 to -6). Here we show that binding of PA700 to the 20 S proteasome is mediated by the COOH termini of two (Rpt2 and Rpt5) of the six Rpt subunits that constitute the interaction surface between the subcomplexes. COOH-terminal peptides of either Rpt2 or Rpt5 bind to the 20 S proteasome and activate hydrolysis of short peptide substrates. Simultaneous binding of both COOH-terminal peptides had additive effects on peptide substrate hydrolysis, suggesting that they bind to distinct sites on the proteasome. In contrast, only the Rpt5 peptide activated hydrolysis of protein substrates. Nevertheless, the COOH-terminal peptide of Rpt2 greatly enhanced this effect, suggesting that proteasome activation is a multistate process. Rpt2 and Rpt5 COOH-terminal peptides cross-linked to different but specific subunits of the 20 S proteasome. These results reveal critical roles of COOH termini of Rpt subunits of PA700 in the assembly and activation of eukaryotic 26 S proteasome. Moreover, they support a model in which Rpt subunits bind to dedicated sites on the proteasome and play specific, nonequivalent roles in the asymmetric assembly and activation of the 26 S proteasome.
Figures
References
-
- Coux, O., Tanaka, K., and Goldberg, A. L. (1996) Annu. Rev. Biochem. 65 801-847 - PubMed
-
- Voges, D., Zwickl, P., and Baumeister, W. (1999) Annu. Rev. Biochem. 68 1015-1068 - PubMed
-
- Pickart, C. M., and Cohen, R. E. (2004) Nat. Rev. Mol. Cell Biol. 5 177-187 - PubMed
-
- Gillette, T. G., and DeMartino, G. N. (2007) Cell 129 659-662 - PubMed
-
- DeMartino, G. N., and Slaughter, C. A. (1999) J. Biol. Chem. 274 22123-22126 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
