

Characterization of orchardgrass p23, a flowering plant Hsp90 cohort protein
- PMID: 18800239
- PMCID: PMC2728258
- DOI: 10.1007/s12192-008-0077-6
Characterization of orchardgrass p23, a flowering plant Hsp90 cohort protein
Abstract
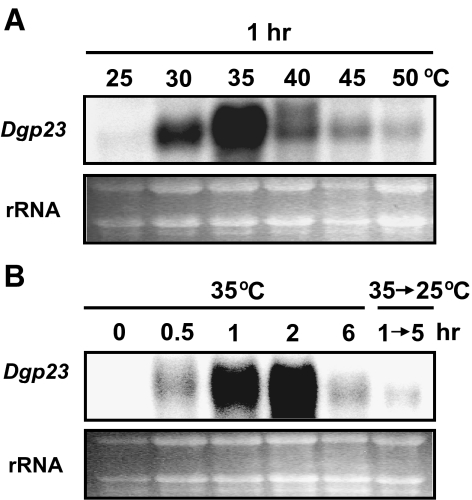
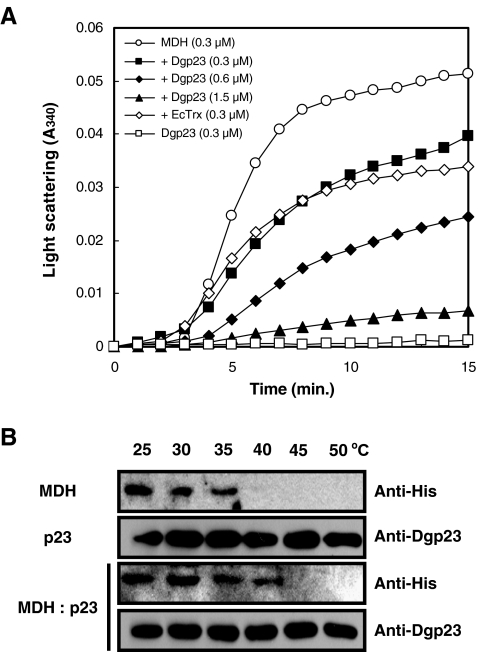
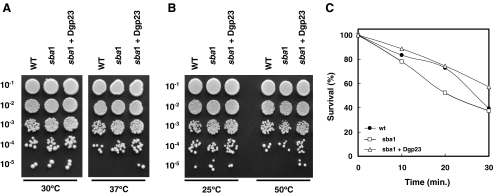
p23 is a heat shock protein 90 (Hsp90) co-chaperone and stabilizes the Hsp90 heterocomplex in mammals and yeast. In this study, we isolated a complementary DNA (cDNA) encoding p23 from orchardgrass (Dgp23) and characterized its functional roles under conditions of thermal stress. Dgp23 is a 911 bp cDNA with an open reading frame predicted to encode a 180 amino acid protein. Northern analysis showed that expression of Dgp23 transcripts was heat inducible. Dgp23 has a well-conserved p23 domain and interacted with an orchardgrass Hsp90 homolog in vivo, like mammalian and yeast p23 homologs. Recombinant Dgp23 is a small acidic protein with a molecular mass of approximately 27 kDa and pI 4.3. Dgp23 was also shown to function as a chaperone protein by suppression of malate dehydrogenase thermal aggregation. Differential scanning calorimetry thermograms indicated that Dgp23 is a heat-stable protein, capable of increasing the T (m) of lysozyme. Moreover, overexpression of Dgp23 in a yeast p23 homolog deletion strain, Deltasba1, increased cell viability. These results suggest that Dgp23 plays a role in thermal stress-tolerance and functions as a co-chaperone of Hsp90 and as a chaperone.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
