

Rapid enhancement of two-step wiring plasticity by estrogen and NMDA receptor activity
- PMID: 18801922
- PMCID: PMC2567160
- DOI: 10.1073/pnas.0801581105
Rapid enhancement of two-step wiring plasticity by estrogen and NMDA receptor activity
Erratum in
- Proc Natl Acad Sci U S A. 2008 Dec 16;105(50):20045.. Woolfrey, Kevin [corrected to Woolfrey, Kevin M]
Abstract
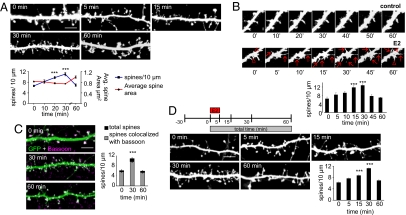
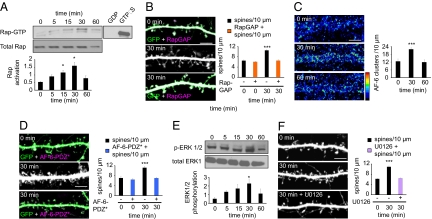
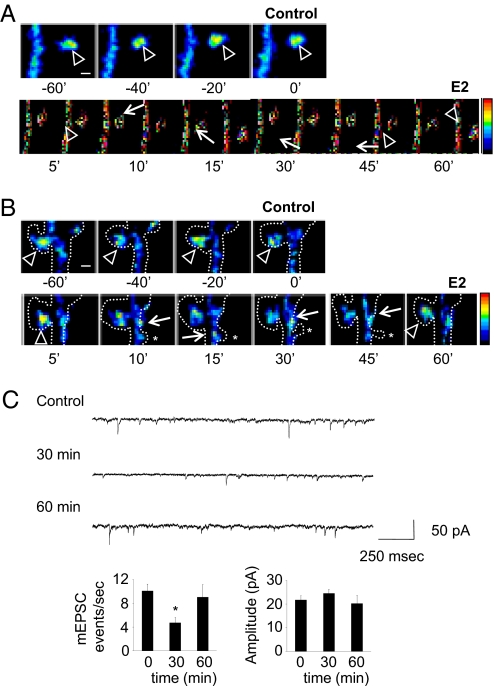
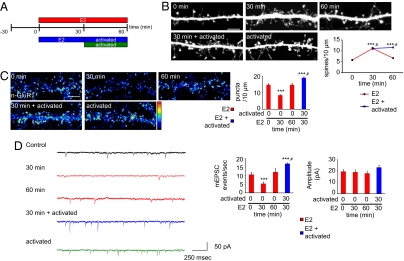
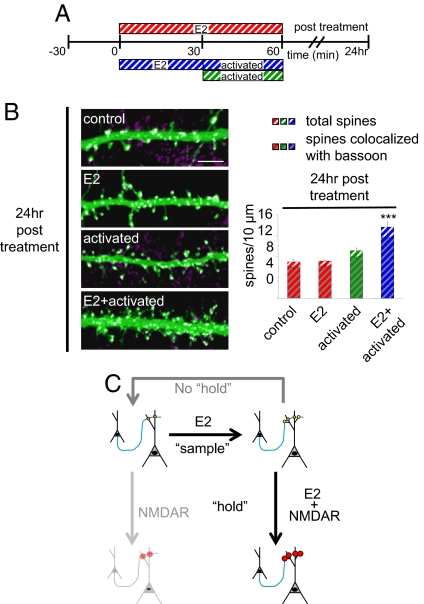
Cortical information storage requires combined changes in connectivity and synaptic strength between neurons, but the signaling mechanisms underlying this two-step wiring plasticity are unknown. Because acute 17beta-estradiol (E2) modulates cortical memory, we examined its effects on spine morphogenesis, AMPA receptor trafficking, and GTPase signaling in cortical neurons. Acute E2 application resulted in a rapid, transient increase in spine density, accompanied by temporary formation of silent synapses through reduced surface GluR1. These rapid effects of E2 were dependent on a Rap/AF-6/ERK1/2 pathway. Intriguingly, NMDA receptor (NMDAR) activation after E2 treatment potentiated silent synapses and elevated spine density for as long as 24 h. Hence, we show that E2 transiently increases neuronal connectivity by inducing dynamic nascent spines that "sample" the surrounding neuropil and that subsequent NMDAR activity is sufficient to stabilize or "hold" E2-mediated effects. This work describes a form of two-step wiring plasticity relevant for cortical memory and identifies targets that may facilitate recovery from brain injuries.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Chklovskii DB, Mel BW, Svoboda K. Cortical rewiring and information storage. Nature. 2004;431:782–788. - PubMed
-
- Holtmaat AJ, et al. Transient and persistent dendritic spines in the neocortex in vivo. Neuron. 2005;45:279–291. - PubMed
-
- Kasai H, Matsuzaki M, Noguchi J, Yasumatsu N, Nakahara H. Structure-stability-function relationships of dendritic spines. Trends Neurosci. 2003;26:360–368. - PubMed
-
- Rumpel S, LeDoux J, Zador A, Malinow R. Postsynaptic receptor trafficking underlying a form of associative learning. Science. 2005;308:83–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
