

Protein kinase C theta regulates stability of the peripheral adhesion ring junction and contributes to the sensitivity of target cell lysis by CTL
- PMID: 18802085
- PMCID: PMC2748977
- DOI: 10.4049/jimmunol.181.7.4815
Protein kinase C theta regulates stability of the peripheral adhesion ring junction and contributes to the sensitivity of target cell lysis by CTL
Abstract
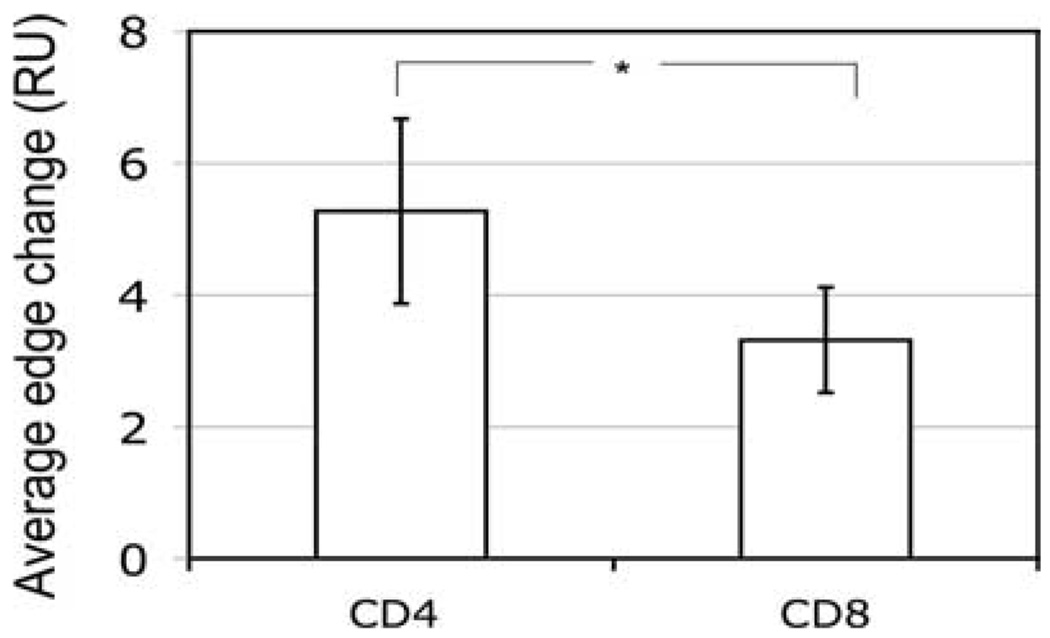
Destruction of virus-infected cells by CTL is an extremely sensitive and efficient process. Our previous data suggest that LFA-1-ICAM-1 interactions in the peripheral supramolecular activation cluster (pSMAC) of the immunological synapse mediate formation of a tight adhesion junction that might contribute to the sensitivity of target cell lysis by CTL. Herein, we compared more (CD8(+)) and less (CD4(+)) effective CTL to understand the molecular events that promote efficient target cell lysis. We found that abrogation of the pSMAC formation significantly impaired the ability of CD8(+) but not CD4(+) CTL to lyse target cells despite having no effect of the amount of released granules by both CD8(+) and CD4(+) CTL. Consistent with this, CD4(+) CTL break their synapses more often than do CD8(+) CTL, which leads to the escape of the cytolytic molecules from the interface. CD4(+) CTL treatment with a protein kinase Ctheta inhibitor increases synapse stability and sensitivity of specific target cell lysis. Thus, formation of a stable pSMAC, which is partially controlled by protein kinase Ctheta, functions to confine the released lytic molecules at the synaptic interface and to enhance the effectiveness of target cell lysis.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures









References
-
- Mentzer SJ, Smith BR, Barbosa JA, Crimmins MA, Herrmann SH, Burakoff SJ. CTL adhesion and antigen recognition are discrete steps in the human CTL-target cell interaction. J. Immunol. 1987;138:1325–1330. - PubMed
-
- Shaw S, Luce GE, Quinones R, Gress RE, Springer TA, Sanders ME. Two antigen-independent adhesion pathways used by human cytotoxic T-cell clones. Nature. 1986;323:262–264. - PubMed
-
- Spitz H, van Schooten W, Keizer H, van Seventer G, van de Rijn M, Terhorst C, de Vries J-E. Alloantigen recognition is preceded by nonspecific adhesion of cytotoxic T cells and target cells. Science. 1986;232:403–405. - PubMed
-
- Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML. The immunological synapse: a molecular machine controlling T cell activation. Science. 1999;285:221–227. - PubMed
-
- Monks C, Freiberg B, Kupfer H, Sciaky N, Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 1998;395:82–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
