

Review
doi: 10.1038/nri2416.
Lineage fate and intense debate: myths, models and mechanisms of CD4- versus CD8-lineage choice
Affiliations
- PMID: 18802443
- PMCID: PMC2760737
- DOI: 10.1038/nri2416
Item in Clipboard
Review
Lineage fate and intense debate: myths, models and mechanisms of CD4- versus CD8-lineage choice
Nat Rev Immunol.
2008 Oct.
Abstract
Following successful gene rearrangement at alphabeta T-cell receptor (TCR) loci, developing thymocytes express both CD4 and CD8 co-receptors and undergo a life-or-death selection event, which is known as positive selection, to identify cells that express TCRs with potentially useful ligand specificities. Positively selected thymocytes must then differentiate into either CD4(+) helper T cells or CD8(+) cytotoxic T cells, a crucial decision known as CD4/CD8-lineage choice. In this Review, we summarize recent advances in our understanding of the cellular and molecular events involved in lineage-fate decision and discuss them in the context of the major models of CD4/CD8-lineage choice.
Figures
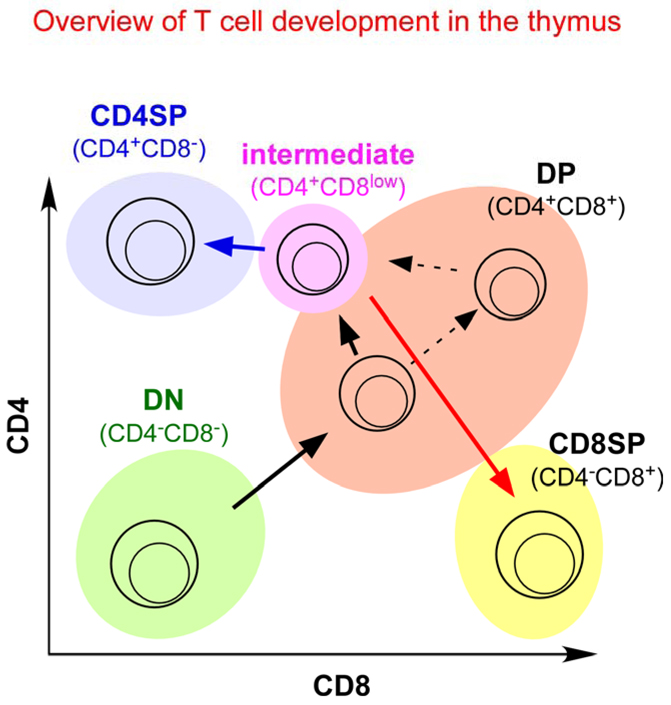
Thymocyte subpopulations can be identified by cell-surface co-receptor expression. Double negative (CD4−CD8−, DN) cells, which express neither CD4 nor CD8, are the most immature cells in the thymus. DN cells differentiate into double positive (CD4+CD8+, DP) thymocytes, which are the first cells to express a functional αβ T-cell receptor (TCR). DP thymocytes that express potentially useful TCR specificities are signalled by the TCR to undergo positive selection and to become intermediate (CD4+CD8low) cells, which then differentiate into either CD4 single positive (CD4+CD8−, CD4 SP) or CD8 SP (CD4−CD8+) mature thymocytes. Depending on the timing of their expression of a functional αβTCR, DP thymocytes can be signalled to undergo positive selection either when they express low levels of both co-receptors or when they express high levels of both co-receptors.
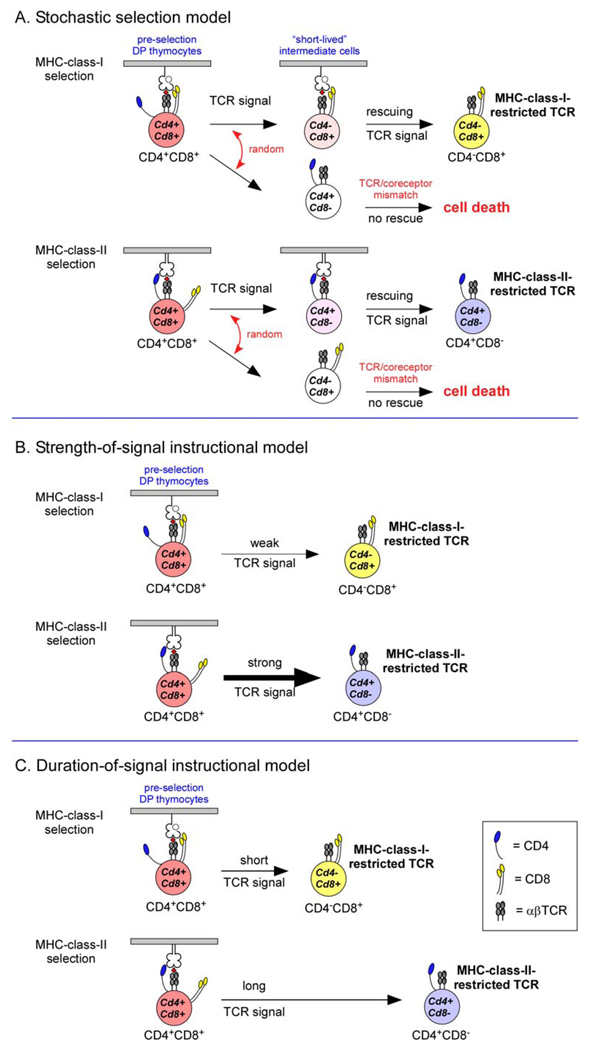
A. The stochastic selection model postulates that positive selecting T-cell receptor (TCR) signals randomly terminate expression of one or the other co-receptor molecule, resulting in the generation of “short-lived” intermediate cells, which will undergo programmed cell death unless rescued by a second TCR signal. Because the TCR-mediated rescue signal requires TCRs and co-receptors that are matched, 50% of positively selected thymocytes will fail to survive and mature. B. The strength-of-signal instructional model postulates that strong TCR signals terminate Cd8 transcription whereas weak TCR signals terminate Cd4 transcription. Signalling by CD4 and MHC-class-II-restricted TCRs is strong resulting in mature CD4+ T cells, whereas signalling by CD8 and MHC-class-I-restricted TCRs is weak resulting in mature CD8+ T cells. C. The duration-of-signal instructional model postulates that long and/or strong TCR signals terminate Cd8 transcription, whereas short and/or weak TCR signals terminate Cd4 transcription. MHC-class-I-restricted and MHC-class-II-restricted TCR signals are proposed to differ in both duration and intensity.
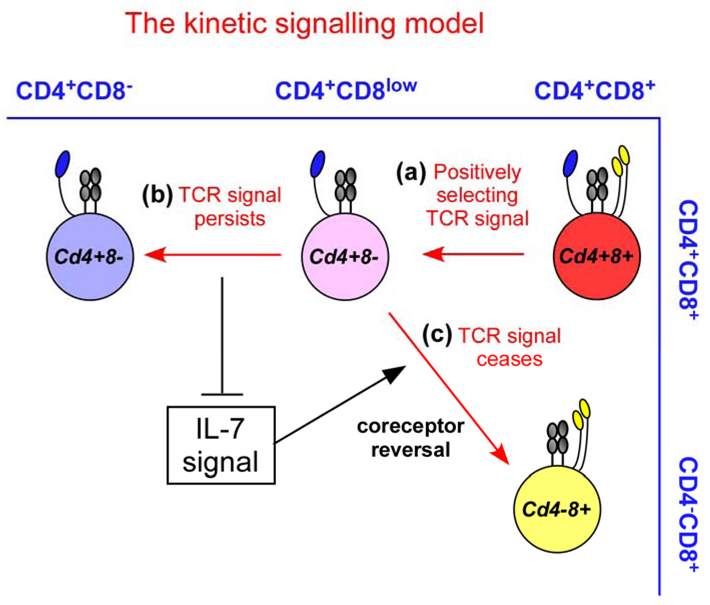
Regardless of the specificity of their T-cell receptor (TCR), positively selecting TCR signals induce double-positive (DP) thymocytes that are transcriptionally Cd4+Cd8+ to terminate Cd8 gene expression and to convert into Cd4+Cd8−intermediate thymocytes. Because of absent Cd8 gene transcription, Cd4+Cd8− intermediate thymocytes appear phenotypically as CD4+CD8low cells (step a), and these are the cells in which lineage choice is made. Persistence of TCR signalling in Cd4+Cd8− intermediate thymocytes blocks interleukin-7 (IL-7) signalling and induces differentiation into mature CD4+ T cells (step b). Cessation or disruption of TCR signalling in Cd4+Cd8− permits IL-7 signalling, which induces Cd4+Cd8− intermediate thymocytes to undergo co-receptor reversal to become Cd4−Cd8+ and to differentiate into CD8+ T cells (step c).
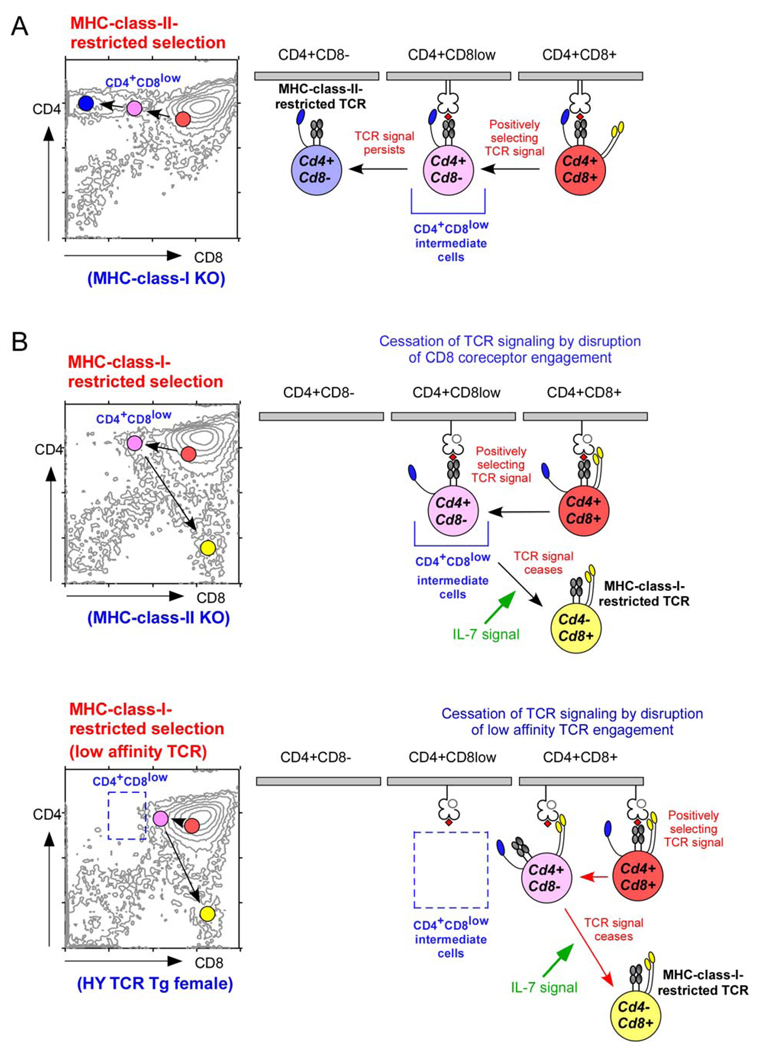
Regardless of the MHC-restriction specificity of the T-cell receptor (TCR), positively selecting TCR signals convert double-positive (DP) thymocytes into Cd4+Cd8− intermediate thymocytes in which lineage choice is made. CD4/CD8 lineage direction is then dependent on whether positively selecting TCR signals persist or cease. A. MHC-class-II-restricted TCR signalling is independent of CD8 expression and, therefore, persists in Cd4+Cd8− intermediate thymocytes. Persistent TCR signalling induces intermediate thymocytes to differentiate into mature CD4+ T cells. B. MHC-class-I-restricted TCR signalling is dependent on CD8 expression and, therefore, ceases in Cd4+Cd8− intermediate thymocytes. Cessation of TCR signalling permits interleukin-7 (IL-7) signalling, which induces intermediate thymocytes to undergo co-receptor reversal and to differentiate into mature CD8+ T cells. The appearance of Cd4+Cd8− intermediate thymocytes at the point that TCR signalling ceases varies according to the ligand affinity of individual TCRs. Signalling by high affinity MHC-class-I-restricted TCRs (upper panel) can persist in Cd4+Cd8− intermediate thymocytes until intermediate thymocytes have lost sufficient CD8 to become CD4+CD8low cells. However, signalling by low affinity MHC-class-I-restricted TCRs (lower panel) is dependent on high CD8 expression and is disrupted by small reductions in cell-surface CD8 levels, when intermediate thymocytes still appear as CD4+CD8+ cells.
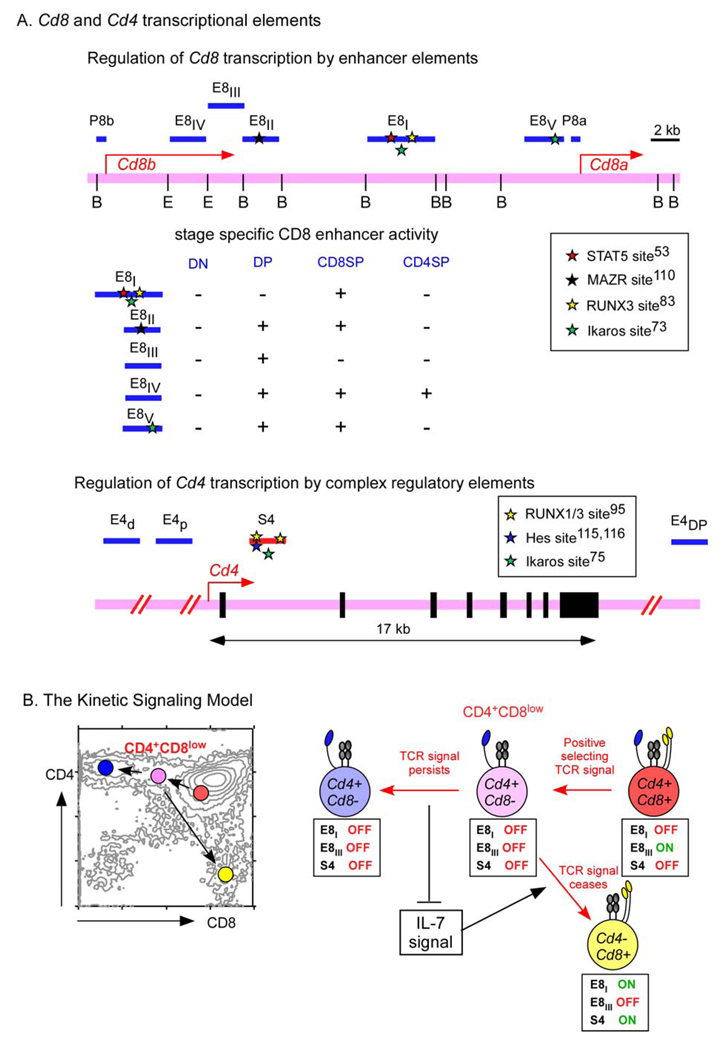
A. Cell-specific expression of the Cd8a gene is controlled by stage-specific enhancer elements, of which five are known (E8I-E8V). Nuclear factors that bind to these regions include Ikaros , RUNX1 (runt-related transcription factor 1), , RUNX3, , , MAZR (MAZ-related factor) and STAT5 (signal transducer and activator of transcription 5). In contrast to Cd8 gene expression, tissue specific expression of the Cd4 gene is not accomplished by Cd4 enhancer elements (E4)–, but is mainly controlled by activation of a silencer element (S4) that is located in the first intron, , , . Nuclear factors that bind to the CD4 silencer element include RUNX, MYB and HES1, , , . B. Changes in co-receptor transcription during positive selection and lineage choice according to the kinetic signalling model. CD8 expression on pre-selection double-positive (DP) thymocytes is driven in part by the E8III Cd8 enhancer, which is turned off by T-cell receptor (TCR)-mediated positive selection signalling, . In CD4+CD8low intermediate thymocytes, persistent TCR-mediated positive selection signalling inhibits Cd8 gene expression and inhibits Cd4 silencer activity, so that intermediate cells differentiate into CD4+ T cells. However, cessation of TCR-mediated positive selection signalling in intermediate thymocytes results in re-initiation of Cd8 gene expression, at least in part, by induction of E8I Cd8 enhancer activity, which is responsive to interleukin-7 receptor (IL-7R) signalling.
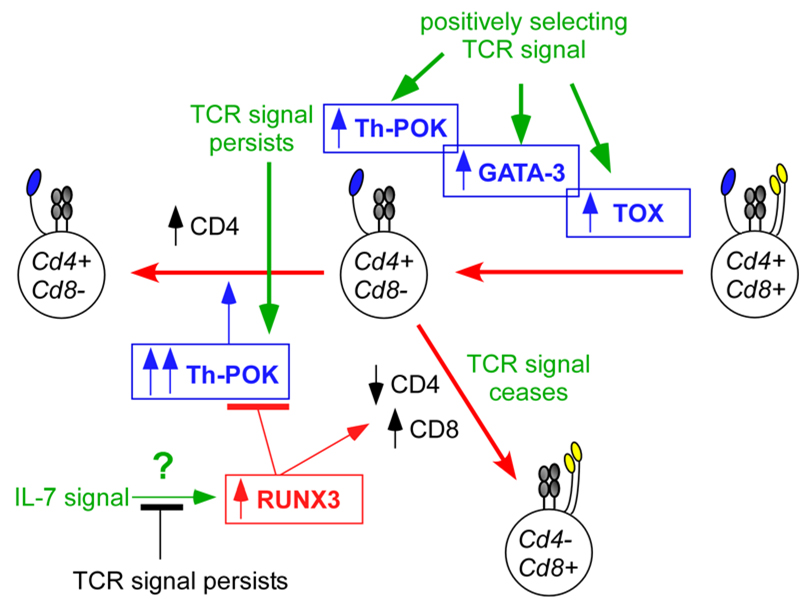
Environmental cues that influence CD4/CD8 lineage choice must ultimately be translated by developing thymocytes into molecular events mediated by nuclear factors that differentially affect co-receptor gene expression. Here, we consider the interactions among four different transcription factors: Th-POK (T-helper-inducing POZ/Kruppel-like factor), RUNX3 (runt-related transcription factor 3), TOX (thymus high-mobility group box protein) and GATA3 (GATA-binding protein 3). Three of these factors are important for CD4+ T-cell differentiation (Th-POK, TOX and GATA3), and only one (RUNX3) is known to be important for CD8+ T-cell differentiation. During positive selection, T-cell receptor (TCR) signals upregulate TOX, GATA3 and Th-POK. TOX upregulation is necessary for TCR-signalled DP thymocytes to phenotypically become CD4+CD8low intermediate thymocytes. GATA3 upregulation is important for the differentiation of CD4+CD8low thymocytes into CD4+ T cells, . And Th-POK expression in TCR-signalled thymocytes, which is significantly upregulated in CD4+CD8low intermediate thymocytes by persistent TCR signalling, is required for CD4-lineage commitment, , and for preventing Cd4 gene silencing by RUNX proteins. It is not yet known what environmental signal upregulates RUNX3 expression, but it is hypothesized that its expression may be upregulated by interleukin-7 receptor (IL-7R) signalling. In any event, RUNX3 performs three important functions that promote the differentiation of intermediate thymocytes into CD8+ T cells: first, RUNX3 binds to the Cd4 silencer element and silences Cd4 gene expression; second, RUNX3 binds to the E8I Cd8 enhancer element and re-initiates Cd8 gene expression, and third RUNX3 silences Th-POK gene expression.
References
-
- Chong MM, et al. Suppressor of cytokine signaling-1 is a critical regulator of interleukin-7-dependent CD8+ T cell differentiation. Immunity. 2003;18:475–487. - PubMed
-
- Doyle C, Strominger JL. Interaction between CD4 and class II MHC molecules mediates cell adhesion. Nature. 1987;330:256–259. - PubMed
-
- Norment AM, Salter RD, Parham P, Engelhard VH, Littman DR. Cell-cell adhesion mediated by CD8 and MHC class I molecules. Nature. 1988;336:79–81. - PubMed
-
- Shaw AS, et al. The lck tyrosine protein kinase interacts with the cytoplasmic tail of the CD4 glycoprotein through its unique amino-terminal domain. Cell. 1989;59:627–636. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
