

Processive kinesins require loose mechanical coupling for efficient collective motility
- PMID: 18802450
- PMCID: PMC2581844
- DOI: 10.1038/embor.2008.169
Processive kinesins require loose mechanical coupling for efficient collective motility
Abstract
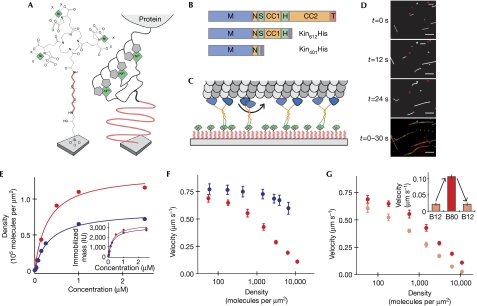
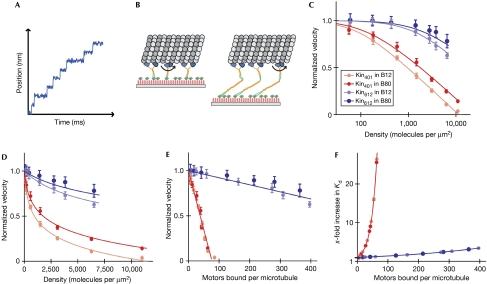
Processive motor proteins are stochastic steppers that perform actual mechanical steps for only a minor fraction of the time they are bound to the filament track. Motors usually work in teams and therefore the question arises whether the stochasticity of stepping can cause mutual interference when motors are mechanically coupled. We used biocompatible surfaces to immobilize processive kinesin-1 motors at controlled surface densities in a mechanically well-defined way. This helped us to study quantitatively how mechanical coupling between motors affects the efficiency of collective microtubule transport. We found that kinesin-1 constructs that lack most of the non-motor sequence slow each other down when collectively transporting a microtubule, depending on the number of interacting motors. This negative interference observed for a motor ensemble can be explained quantitatively by a mathematical model using the known physical properties of individual molecules of kinesin-1. The non-motor extension of kinesin-1 reduces this mutual interference, indicating that loose mechanical coupling between motors is required for efficient transport by ensembles of processive motors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Berliner E, Mahtani HK, Karki S, Chu LF, Cronan JE Jr, Gelles J (1994) Microtubule movement by a biotinylated kinesin bound to streptavidin-coated surface. J Biol Chem 269: 8610–8615 - PubMed
-
- Berliner E, Young EC, Anderson K, Mahtani HK, Gelles J (1995) Failure of a single-headed kinesin to track parallel to microtubule protofilaments. Nature 373: 718–721 - PubMed
-
- Bieling P, Laan L, Schek H, Munteanu EL, Sandblad L, Dogterom M, Brunner D, Surrey T (2007) Reconstitution of a microtubule plus-end tracking system in vitro. Nature 450: 1100–1105 - PubMed
-
- Carter NJ, Cross RA (2005) Mechanics of the kinesin step. Nature 435: 308–312 - PubMed
-
- Case RB, Pierce DW, Hom-Booher N, Hart CL, Vale RD (1997) The directional preference of kinesin motors is specified by an element outside of the motor catalytic domain. Cell 90: 959–966 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
