

A mouse model of harlequin ichthyosis delineates a key role for Abca12 in lipid homeostasis
- PMID: 18802465
- PMCID: PMC2529452
- DOI: 10.1371/journal.pgen.1000192
A mouse model of harlequin ichthyosis delineates a key role for Abca12 in lipid homeostasis
Erratum in
- PLoS Genet. 2008 Oct;4(10). doi: 10.1371/annotation/02a793a0-160f-46b9-abaa-4a3d3eecdde2. Slattery, Keith [corrected to Satterley, Keith] doi: 10.1371/annotation/02a793a0-160f-46b9-abaa-4a3d3eecdde2
Abstract
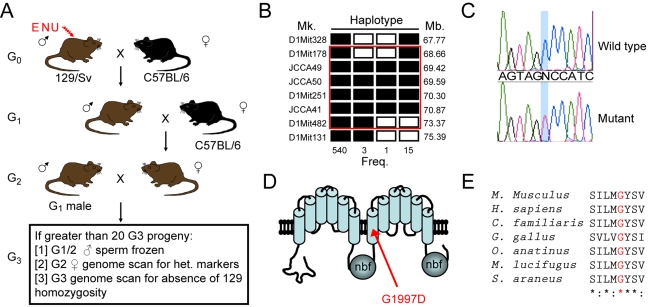
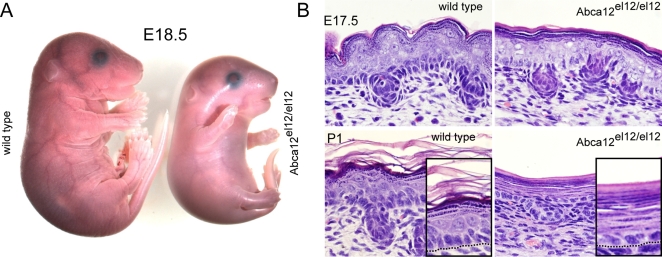
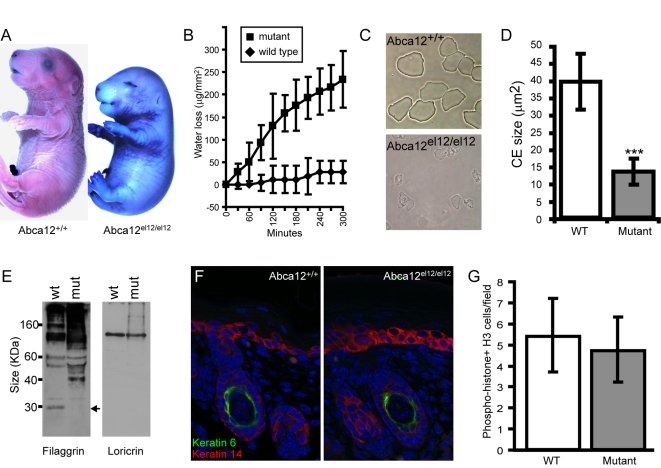
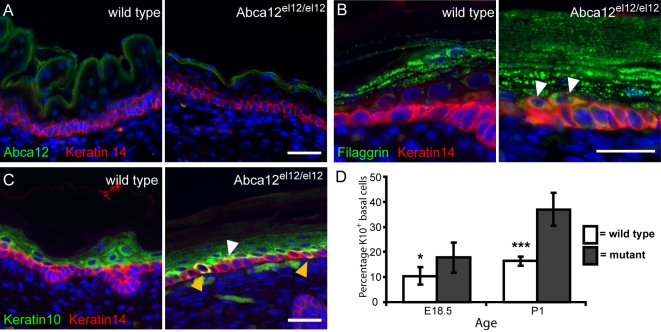
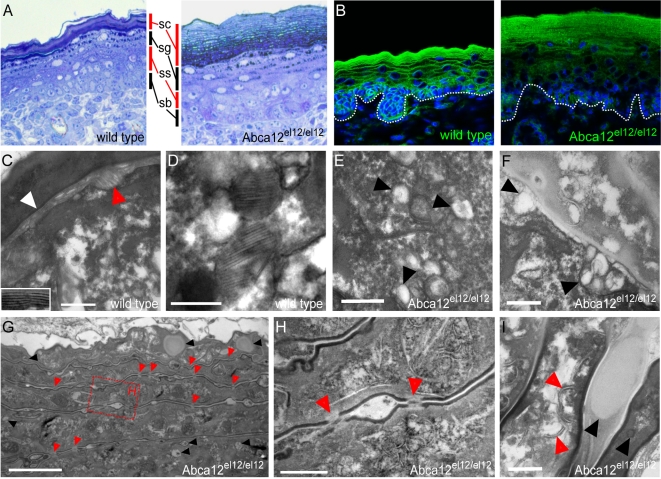
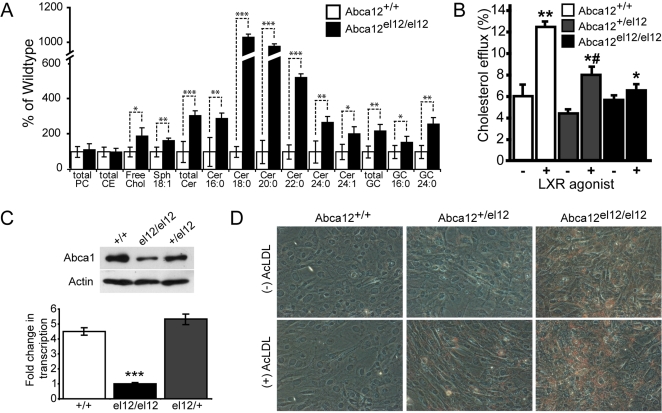
Harlequin Ichthyosis (HI) is a severe and often lethal hyperkeratotic skin disease caused by mutations in the ABCA12 transport protein. In keratinocytes, ABCA12 is thought to regulate the transfer of lipids into small intracellular trafficking vesicles known as lamellar bodies. However, the nature and scope of this regulation remains unclear. As part of an original recessive mouse ENU mutagenesis screen, we have identified and characterised an animal model of HI and showed that it displays many of the hallmarks of the disease including hyperkeratosis, loss of barrier function, and defects in lipid homeostasis. We have used this model to follow disease progression in utero and present evidence that loss of Abca12 function leads to premature differentiation of basal keratinocytes. A comprehensive analysis of lipid levels in mutant epidermis demonstrated profound defects in lipid homeostasis, illustrating for the first time the extent to which Abca12 plays a pivotal role in maintaining lipid balance in the skin. To further investigate the scope of Abca12's activity, we have utilised cells from the mutant mouse to ascribe direct transport functions to the protein and, in doing so, we demonstrate activities independent of its role in lamellar body function. These cells have severely impaired lipid efflux leading to intracellular accumulation of neutral lipids. Furthermore, we identify Abca12 as a mediator of Abca1-regulated cellular cholesterol efflux, a finding that may have significant implications for other diseases of lipid metabolism and homeostasis, including atherosclerosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Singh S, Bhura M, Maheshwari A, Kumar A, Singh CP, et al. Successful treatment of harlequin ichthyosis with acitretin. Int J Dermatol. 2001;40:472–473. - PubMed
-
- Uitto J. The gene family of ABC transporters–novel mutations, new phenotypes. Trends Mol Med. 2005;11:341–343. - PubMed
-
- Elias P, Feingold KR, F M. In: Epidermal lamellar body as a multifunctional secretory organelle; Elias P, Feingold KR, editors. New York: Taylor and Francis; 2006. pp. 261–272.
-
- Wertz PW, Downing DT. Covalently bound omega-hydroxyacylsphingosine in the stratum corneum. Biochim Biophys Acta. 1987;917:108–111. - PubMed
-
- Tybulewicz VL, Tremblay ML, LaMarca ME, Willemsen R, Stubblefield BK, et al. Animal model of Gaucher's disease from targeted disruption of the mouse glucocerebrosidase gene. Nature. 1992;357:407–410. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
