

Pitx2 deletion in pituitary gonadotropes is compatible with gonadal development, puberty, and fertility
- PMID: 18802953
- PMCID: PMC2923441
- DOI: 10.1002/dvg.20398
Pitx2 deletion in pituitary gonadotropes is compatible with gonadal development, puberty, and fertility
Abstract
This report introduces a gonadotrope-specific cre transgenic mouse capable of ablating floxed genes in mature pituitary gonadotropes. Initial analysis of this transgenic line, Tg(Lhb-cre)1Sac, reveals that expression is limited to the pituitary cells that produce luteinizing hormone beta, beginning appropriately at e17.5. Cre activity is detectable by a reporter gene in nearly every LHbeta-producing cell, but the remaining hormone-producing cell types and other organs exhibit little to no activity. We used the Tg(Lhb-cre)1Sac strain to assess the role Pitx2 in gonadotrope function. The gonadotrope-specific Pitx2 knockout mice exhibit normal expression of LHbeta, sexual maturation, and fertility, suggesting that Pitx2 is not required for gonadotrope maintenance or for regulated production of gonadotropins.
Copyright 2008 Wiley-Liss, Inc.
Figures
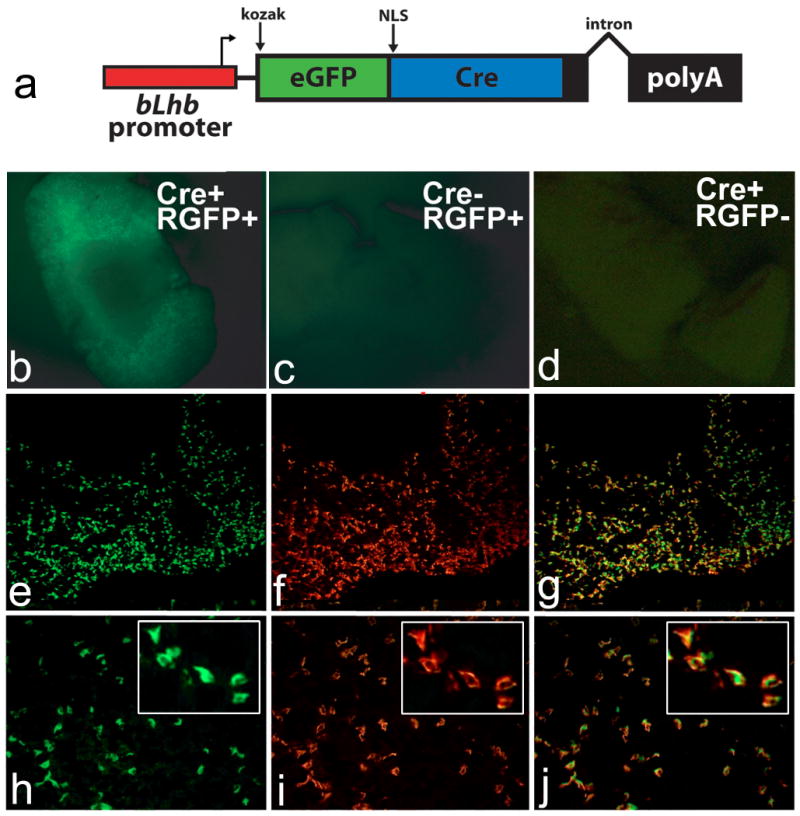
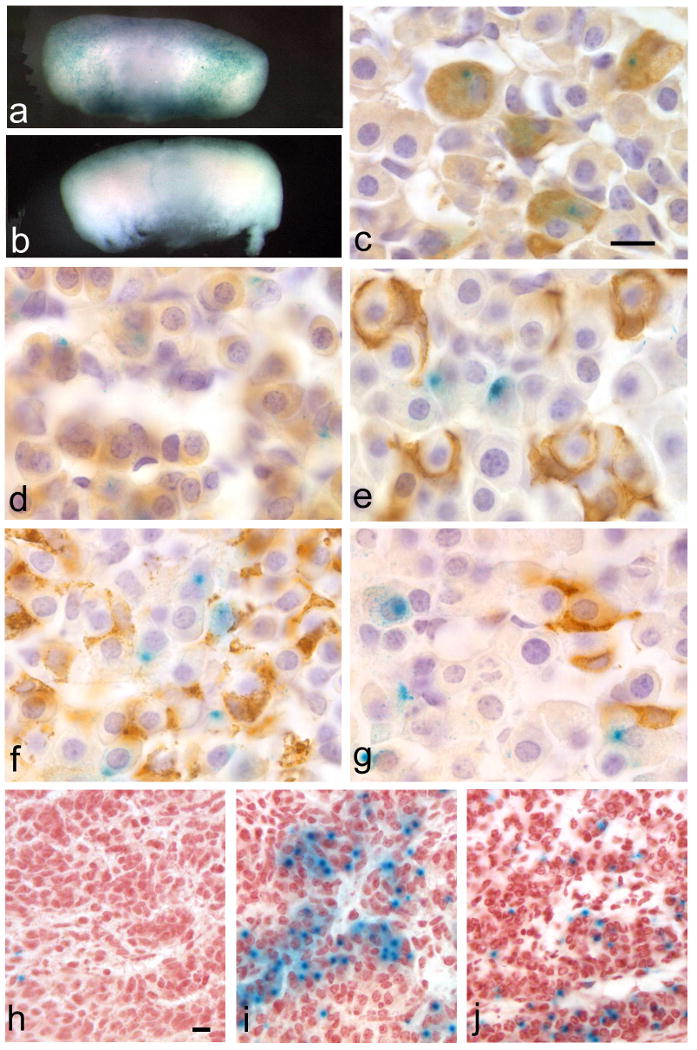
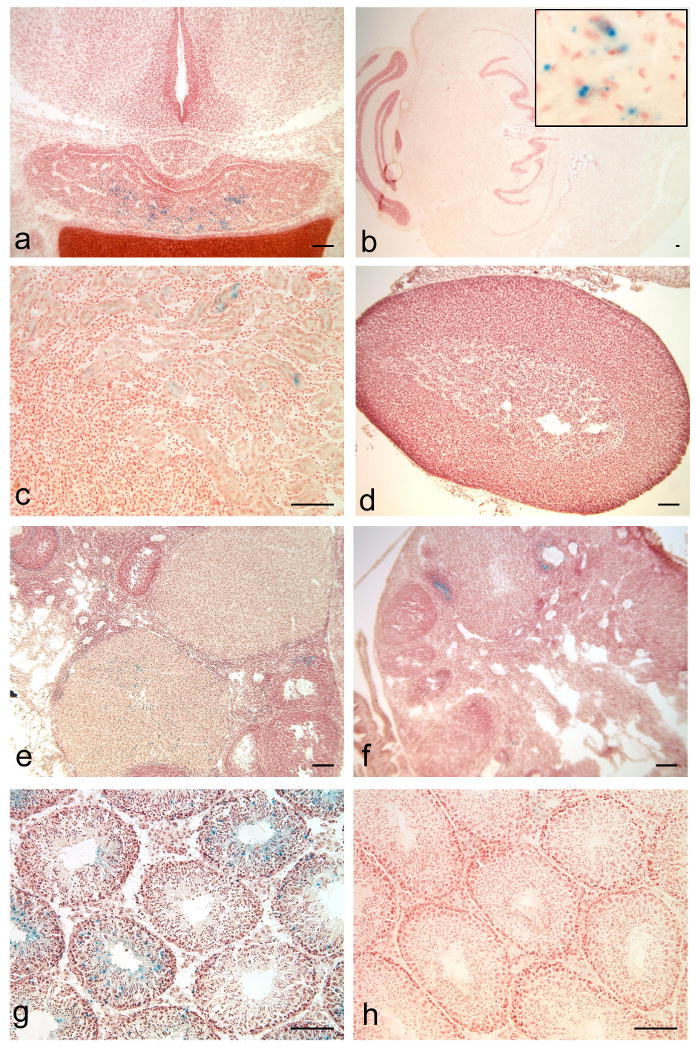

References
-
- Berger P, Kranewitter W, Madersbacher S, Gerth R, Geley S, Dirnhofer S. Eutopic production of human chorionic gonadotropin beta (hCG beta) and luteinizing hormone beta (hLH beta) in the human testis. FEBS Lett. 1994;343:229–233. - PubMed
-
- Bingham NC, Verma-Kurvari S, Parada LF, Parker KL. Development of a steroidogenic factor 1/Cre transgenic mouse line. Genesis. 2006;44:419–424. - PubMed
-
- Brinkmeier ML, Gordon DF, Dowding JM, Saunders TL, Kendall SK, Sarapura VD, Wood WM, Ridgway EC, Camper SA. Cell-specific expression of the mouse glycoprotein hormone alpha-subunit gene requires multiple interacting DNA elements in transgenic mice and cultured cells. Mol Endocrinol. 1998;12:622–633. - PubMed
-
- Burns KH, Matzuk MM. Minireview: genetic models for the study of gonadotropin actions. Endocrinology. 2002;143:2823–2835. - PubMed
-
- Burrows HL, Douglas KR, Seasholtz AF, Camper SA. Genealogy of the Anterior Pituitary Gland: Tracing a Family Tree. Trends Endocrinol Metab. 1999;10:343–352. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
