

Pathogenic missense MAPT mutations differentially modulate tau aggregation propensity at nucleation and extension steps
- PMID: 18803694
- PMCID: PMC2596975
- DOI: 10.1111/j.1471-4159.2008.05692.x
Pathogenic missense MAPT mutations differentially modulate tau aggregation propensity at nucleation and extension steps
Abstract
Mutations in the MAPT gene encoding tau protein lead to neurofibrillary lesion formation, neurodegeneration, and cognitive decline associated with frontotemporal lobar degeneration. While some pathogenic mutations affect MAPT introns, resulting in abnormal splicing patterns, the majority occur in the tau coding sequence leading to single amino acid changes in tau primary structure. Depending on their location within the polypeptide chain, tau missense mutations have been reported to augment aggregation propensity. To determine the mechanisms underlying mutation-associated changes in aggregation behavior, the fibrillization of recombinant pathogenic mutants R5L, G272V, P301L, V337M, and R406W prepared in a full-length four-repeat human tau background was examined in vitro as a function of time and submicromolar tau concentrations using electron microscopy assay methods. Kinetic constants for nucleation and extension phases of aggregation were then estimated by direct measurement and mathematical simulation. Results indicated that the mutants differ from each other and from wild-type tau in their aggregation propensity. G272V and P301L mutations increased the rates of both filament nucleation and extension reactions, whereas R5L and V337M increased only the nucleation phase. R406W did not differ from wild-type in any kinetic parameter. The results show that missense mutations can directly promote tau filament formation at different stages of the aggregation pathway.
Figures
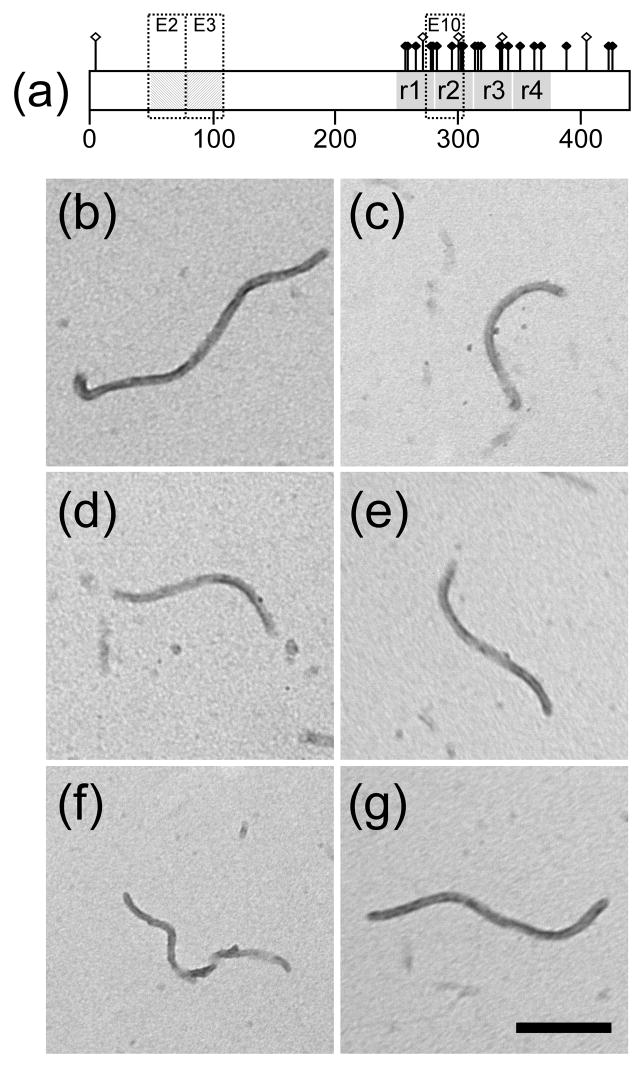
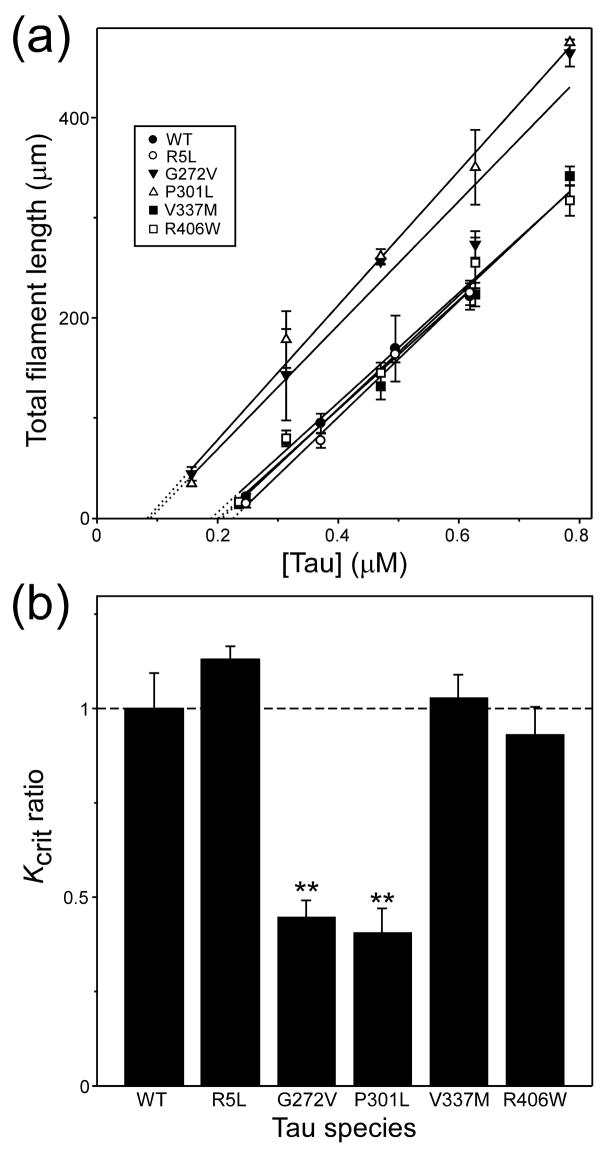
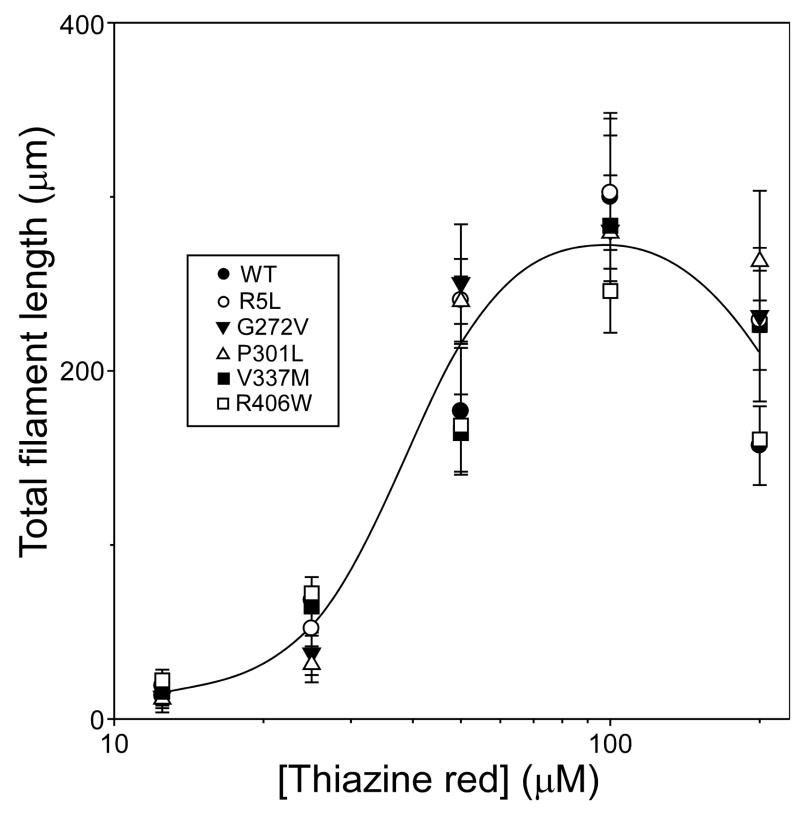

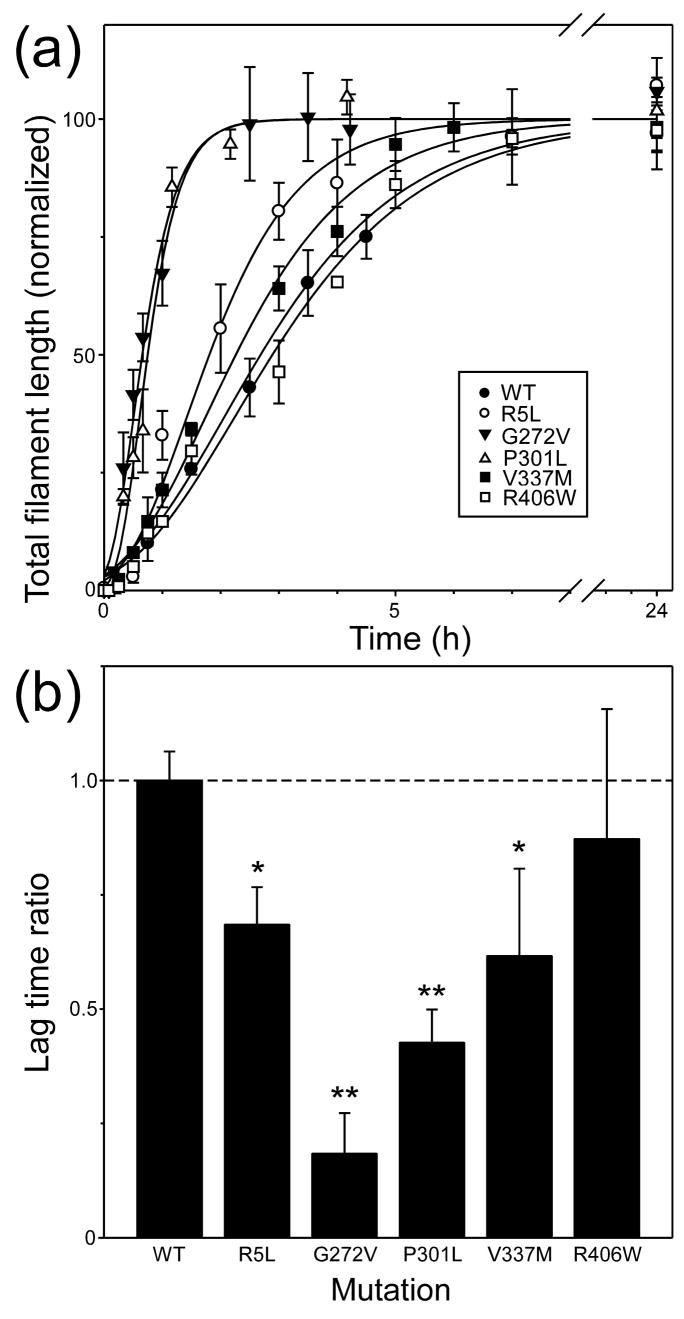


References
-
- Abraha A, Ghoshal N, Gamblin TC, Cryns V, Berry RW, Kuret J, Binder LI. C-terminal inhibition of tau assembly in vitro and in Alzheimer’s disease. J Cell Sci. 2000;113:3737–3745. - PubMed
-
- Ackmann M, Wiech H, Mandelkow E. Nonsaturable binding indicates clustering of tau on the microtubule surface in a paired helical filament-like conformation. The Journal of biological chemistry. 2000;275:30335–30343. - PubMed
-
- Alonso A, Mederlyova A, Novak M, Grundke-Iqbal I, Iqbal K. Promotion of hyperphosphorylation by frontotemporal dementia tau mutations. The Journal of biological chemistry. 2004;279:34873–34881. - PubMed
-
- Barghorn S, Zheng-Fischhofer Q, Ackmann M, Biernat J, von Bergen M, Mandelkow EM, Mandelkow E. Structure, microtubule interactions, and paired helical filament aggregation by tau mutants of frontotemporal dementias. Biochemistry. 2000;39:11714–11721. - PubMed
-
- Biernat J, Gustke N, Drewes G, Mandelkow EM, Mandelkow E. Phosphorylation of Ser262 strongly reduces binding of tau to microtubules: distinction between PHF-like immunoreactivity and microtubule binding. Neuron. 1993;11:153–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
