

A heterospecific leucine zipper tetramer
- PMID: 18804028
- PMCID: PMC7111190
- DOI: 10.1016/j.chembiol.2008.07.008
A heterospecific leucine zipper tetramer
Abstract
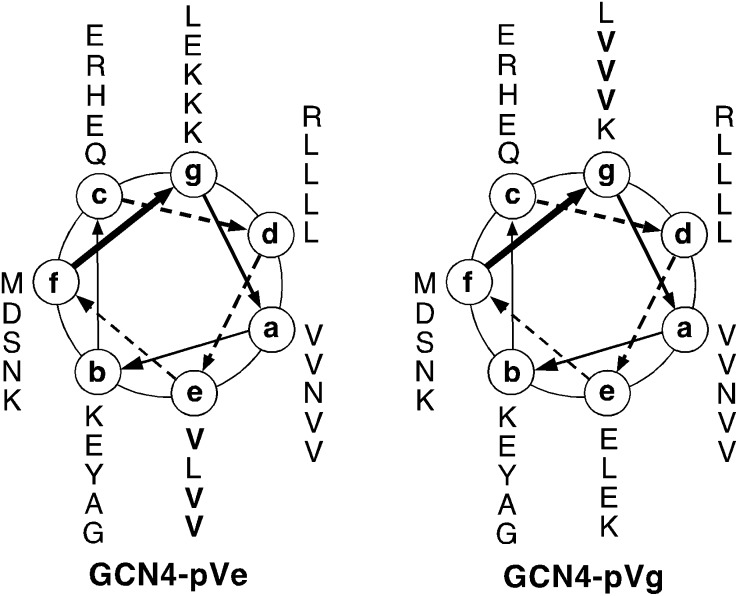
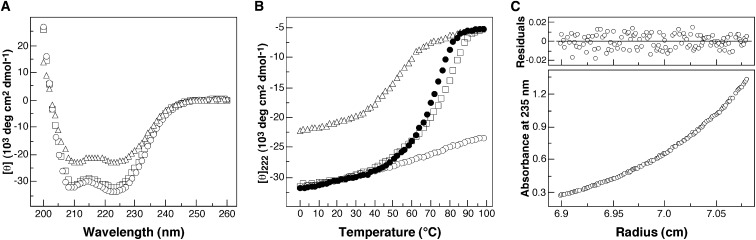
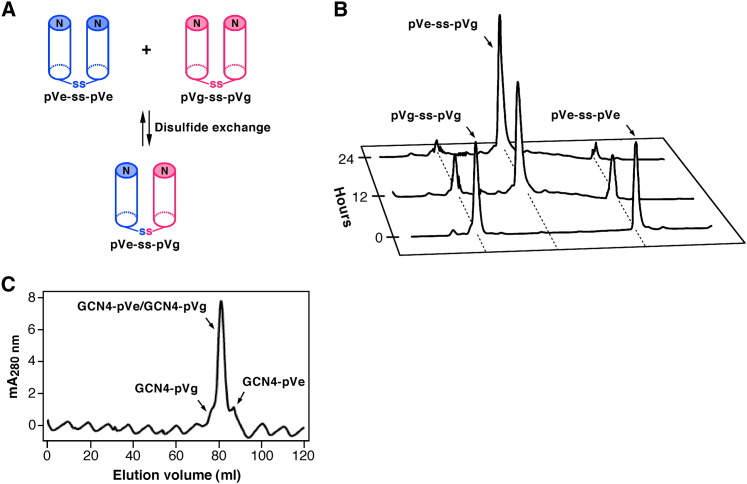
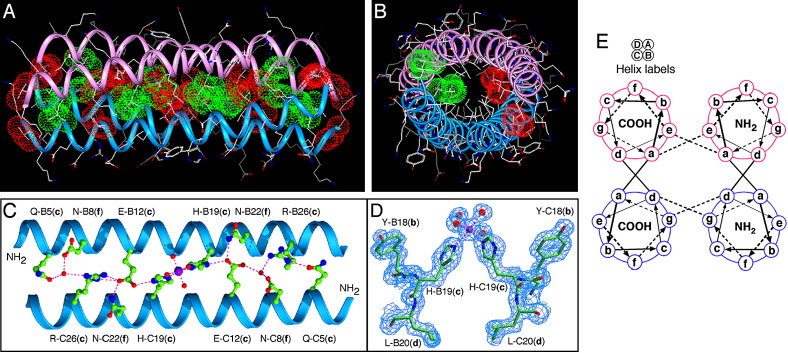
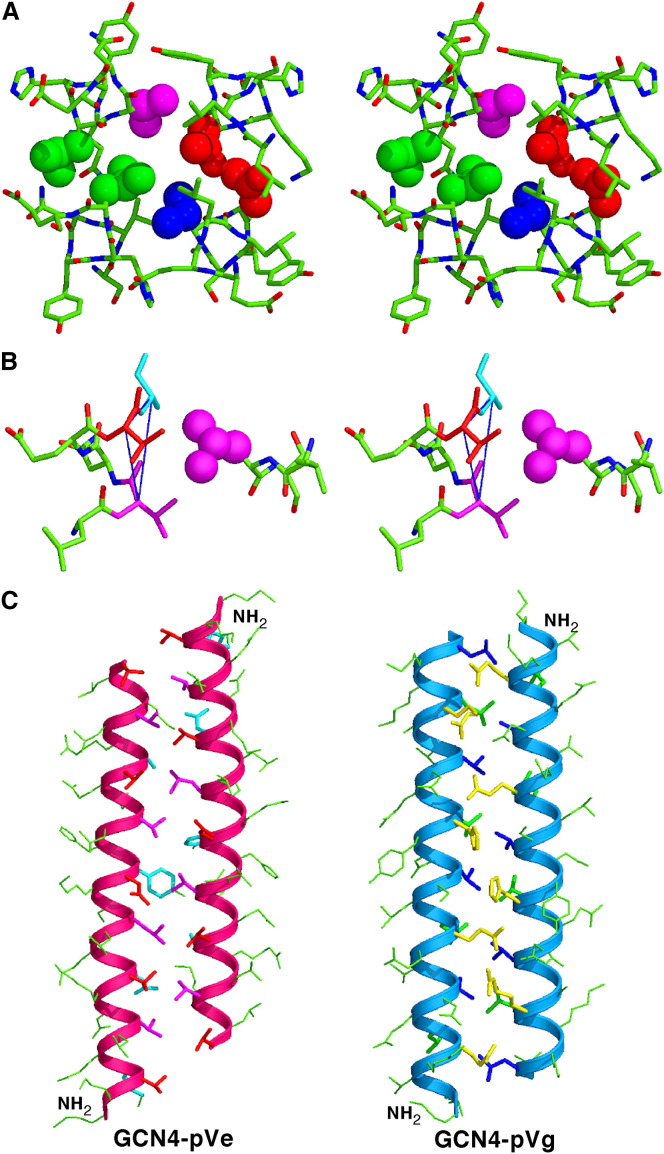
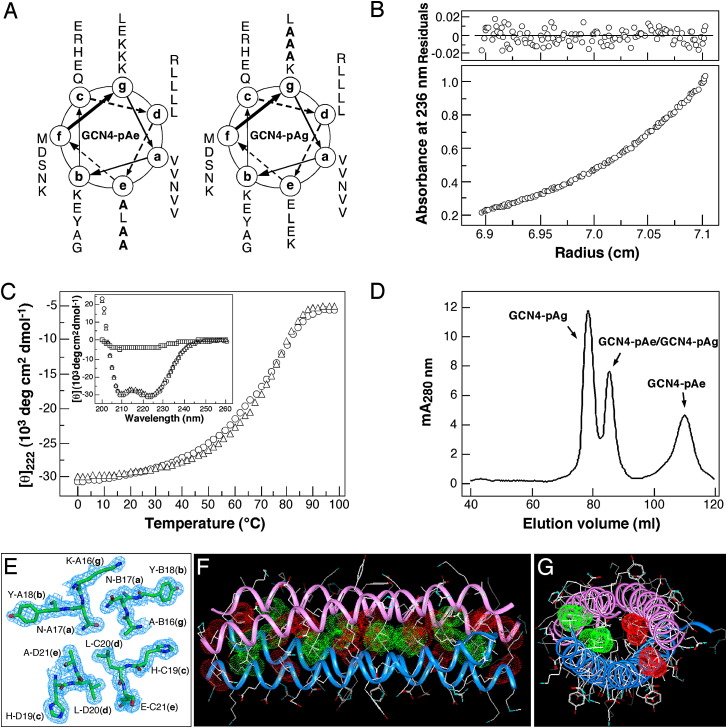
Protein-protein interactions play an essential role in the assembly of the macromolecular complexes that form functional networks and control cellular behavior. Elucidating principles of molecular recognition governing potentially complex interfaces is a challenging goal for structural and systems biology. Extensive studies of alpha-helical coiled coils have provided fundamental insight into the determinants of one seemingly tractable class of oligomeric protein interfaces. We report here that two different valine-containing mutants of the GCN4 leucine zipper that fold individually as four-stranded coiled coils associate preferentially in mixtures to form an antiparallel, heterotetrameric structure. X-ray crystallographic analysis reveals that the coinciding hydrophobic interfaces of the hetero- and homotetramers differ in detail, thereby controlling their partnering and structural specificity. Equilibrium disulfide exchange and thermal denaturation experiments show that the 50-fold preference for heterospecificity is determined by interfacial van der Waals interactions and hydrophobicity. Parallel studies of two alanine-containing variants confirm the above-mentioned interpretation of the basis and mechanism of this heterospecificity. Our results suggest that coiled-coil recognition is an inherently geometric process in which heterotypic interaction specificity derives from a complementarity of both shape and chemistry.
Figures
Similar articles
-
A parallel coiled-coil tetramer with offset helices.Biochemistry. 2006 Dec 26;45(51):15224-31. doi: 10.1021/bi061914m. Epub 2006 Nov 29. Biochemistry. 2006. PMID: 17176044
-
Self-assembly of coiled-coil tetramers in the 1.40 A structure of a leucine-zipper mutant.Protein Sci. 2007 Feb;16(2):323-8. doi: 10.1110/ps.062590807. Epub 2006 Dec 22. Protein Sci. 2007. PMID: 17189475 Free PMC article.
-
Protein destabilization by electrostatic repulsions in the two-stranded alpha-helical coiled-coil/leucine zipper.Protein Sci. 1995 Feb;4(2):237-50. doi: 10.1002/pro.5560040210. Protein Sci. 1995. PMID: 7757012 Free PMC article.
-
A switch between two-, three-, and four-stranded coiled coils in GCN4 leucine zipper mutants.Science. 1993 Nov 26;262(5138):1401-7. doi: 10.1126/science.8248779. Science. 1993. PMID: 8248779
-
Pharmacological interference with protein-protein interactions mediated by coiled-coil motifs.Handb Exp Pharmacol. 2008;(186):461-82. doi: 10.1007/978-3-540-72843-6_19. Handb Exp Pharmacol. 2008. PMID: 18491064 Review.
Cited by
-
The 1.9 A structure of the branched-chain amino-acid transaminase (IlvE) from Mycobacterium tuberculosis.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009 Nov 1;65(Pt 11):1071-7. doi: 10.1107/S1744309109036690. Epub 2009 Oct 13. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2009. PMID: 19923721 Free PMC article.
-
Bind&Bite: covalently stabilized heterodimeric coiled-coil peptides for the site-selective, cysteine-free chemical modification of proteins.RSC Chem Biol. 2023 Aug 8;4(10):794-803. doi: 10.1039/d3cb00122a. eCollection 2023 Oct 4. RSC Chem Biol. 2023. PMID: 37799587 Free PMC article.
-
Protein interaction module-assisted function X (PIMAX) approach to producing challenging proteins including hyperphosphorylated tau and active CDK5/p25 kinase complex.Mol Cell Proteomics. 2015 Jan;14(1):251-62. doi: 10.1074/mcp.O114.044412. Epub 2014 Nov 10. Mol Cell Proteomics. 2015. PMID: 25385071 Free PMC article.
-
The Structure and Topology of α-Helical Coiled Coils.Subcell Biochem. 2017;82:95-129. doi: 10.1007/978-3-319-49674-0_4. Subcell Biochem. 2017. PMID: 28101860 Free PMC article. Review.
References
-
- Acharya A., Ruvinov S.B., Gal J., Moll J.R., Vinson C. A heterodimerizing leucine zipper coiled coil system for examining the specificity of a position interactions: amino acids I, V, L, N, A, and K. Biochemistry. 2002;41:14122–14131. - PubMed
-
- Adamson J.G., Zhou N.E., Hodges R.S. Structure, function and application of the coiled-coil protein folding motif. Curr. Opin. Biotechnol. 1993;4:428–437. - PubMed
-
- Brunger A.T., Adams P.D., Clore G.M., DeLano W.L., Gros P., Grosse-Kunstleve R.W., Jiang J.S., Kuszewski J., Nilges M., Pannu N.S. Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 1998;54:905–921. - PubMed
-
- Bryson J.W., Betz S.F., Lu H.S., Suich D.J., Zhou H.X., O'Neil K.T., DeGrado W.F. Protein design: a hierarchic approach. Science. 1995;270:935–941. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
