

Review
doi: 10.1016/j.devcel.2008.08.012.
The role of autophagy in mammalian development: cell makeover rather than cell death
Affiliations
- PMID: 18804433
- PMCID: PMC2688784
- DOI: 10.1016/j.devcel.2008.08.012
Item in Clipboard
Review
The role of autophagy in mammalian development: cell makeover rather than cell death
Dev Cell.
2008 Sep.
Abstract
Autophagy is important for the degradation of bulk cytoplasm, long-lived proteins, and entire organelles. In lower eukaryotes, autophagy functions as a cell death mechanism or as a stress response during development. However, autophagy's significance in vertebrate development, and the role (if any) of vertebrate-specific factors in its regulation, remains unexplained. Through careful analysis of the current autophagy gene mutant mouse models, we propose that in mammals, autophagy may be involved in specific cytosolic rearrangements needed for proliferation, death, and differentiation during embryogenesis and postnatal development. Thus, autophagy is a process of cytosolic "renovation," crucial in cell fate decisions.
Figures
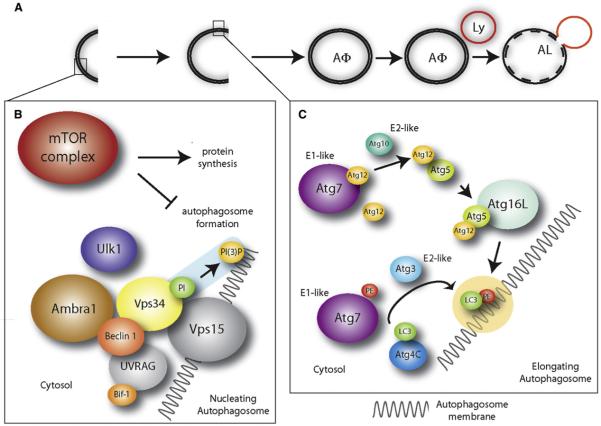
(A) Autophagosomes (AΦ) originate from a precursor structure (phagophore), which grows, closes, and wraps the cytoplasmic components and organelles. Eventually, the autophagosome fuses with the lysosome (Ly), forming a structure termed autophagolysosome or autolysosome (AL). The contents of the autophagosome are then degraded by the lysosomal enzymes. (B) Autophagosome nucleation is driven by phosphatidylinositol (PI) phosphorylation. This process (highlighted in blue) is mediated by a lipid kinase signaling complex (Beclin 1, Vps15, Vps34). Ambra1 promotes Beclin 1/Vps34 interaction, whereas Ulk1 is downstream of mTOR (which is inhibitory for autophagy) and is involved in autophagy induction (see text), although it is not a subunit of the Beclin 1/Vps34 complex. UVRAG and Bif-1 have been described as additional regulators of the Beclin 1/Vps34 complex. (C) Autophagosome elongation is triggered by lipid modification of LC3 (by phosphatidylethanolamine, PE), as highlighted in yellow. This process is mediated, among others, by Atg7 (an E1-like ubiquitin conjugating enzyme), Atg3 (an E2-like ubiquitin conjugating enzyme), and Atg4C, to which LC3 is bound at first. Atg7 also acts in a ubiquitin-like conjugation system involving the E2-like ubiquitin conjugating enzyme Atg10 and Atg12/Atg5 which, at the end of the process, are transferred to Atg16L. The complex Atg12/Atg5/Atg16L mediates LC3-PE binding to the autophagosome membranes.
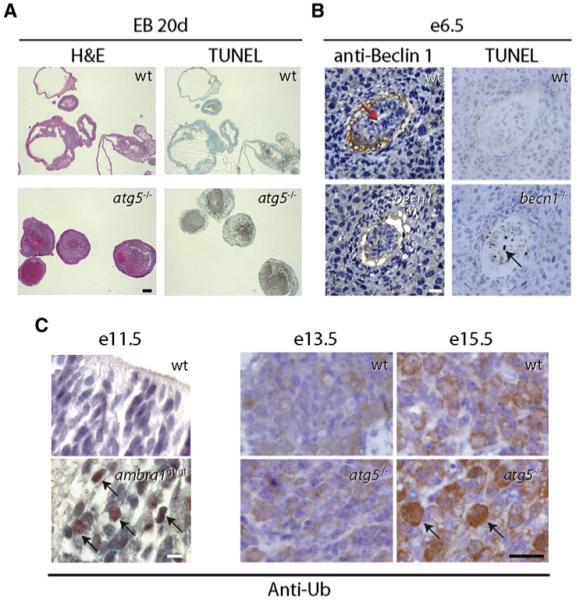
(A) Representative images showing lack of cavity formation and accumulation of TUNEL-positive cells at day 20 (20d) of development of wild-type (wt) and atg5-/- embryoid bodies (EBs) (excerpt from Qu et al., 2007). Similar findings were obtained with becn1-/- embryoid bodies. Scale bar, 200 μm. H&E, hematoxylin and eosin staining; TUNEL, terminal uridine deoxynucleotidyl transferase dUTP nick end labeling for detection of DNA fragmentation (a hallmark of apoptosis). (B) Becn1-/- embryos at e6.5 fail to cavitate (left panels; red arrow labels proamniotic cavity in wt embryo; sections stained with anti-Beclin 1 antibody) and have increased numbers of TUNEL-positive cells (right panels; black arrow). Scale bar, 150 μm. (B.L., unpublished data.) (C) Developing brains lacking ambra1 or atg5 stained positive for ubiquitin (black arrows) at e11.5 and e15.5, respectively (excerpts from Fimia et al., 2007 and Hara et al., 2006). Sections were stained with an antibody directed against ubiquitin (anti-Ub). Scale bars, 10 μm (e11.5) and 25 μm (e13.5/e15.5). Right subpanels reprinted by permission from Macmillan Publishers Ltd: Nature, Hara et al., 441, 885-889, copyright 2006.
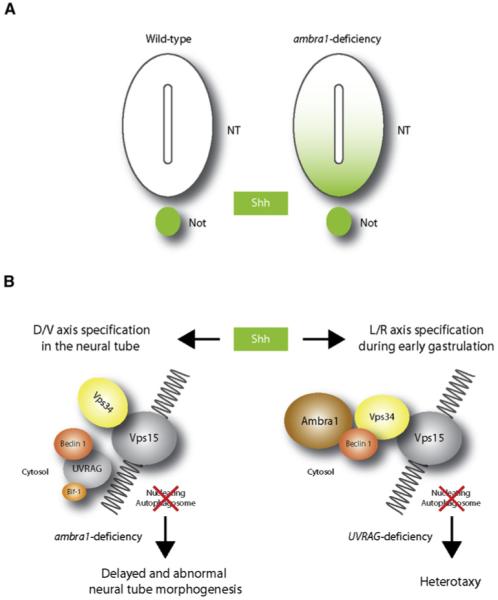
The morphogen sonic hedgehog (Shh) is involved in several aspects of vertebrate development. The model shown is speculative, and autophagy-independent effects may also explain the phenotypes of ambra1, becn1, and UVRAG targeted mutant mice. (A) In wild-type embryos at e9.5, Shh expression is retained within the notochord (Not). At the same developmental stage, ambra1-deficient embryos show Shh expression diffused in the ventral half of the neural tube (NT) and in the floor plate. (B) In the absence of Ambra1, the interaction between Beclin 1 and Vps34 is impaired and autophagy cannot proceed. In addition to the control of Shh signaling during dorso-ventral (D/V) differentiation of the neural tube, autophagy may be involved in regulating the left-right (L/R) axis. In fact, in wild-type embryos, Shh also signals the establishment of the left-right axis in the node cells (see text). In UVRAG mutant humans and Drosophila, the left-right axis symmetry is disturbed, leading to heterotaxy. This could be a consequence of incorrect Shh distribution within the node.
References
-
- Akao Y, Otsuki Y, Kataoka S, Ito Y, Tsujimoto Y. Multiple subcellular localization of bcl-2: detection in nuclear outer membrane, endoplasmic reticulum membrane, and mitochondrial membranes. Cancer Res. 1994;54:2468–2471. - PubMed
-
- Akdemir F, Farkas R, Chen P, Juhasz G, Medved’ova L, Sass M, Wang L, Wang X, Chittaranjan S, Gorski SM, et al. Autophagy occurs upstream or parallel to the apoptosome during histolytic cell death. Development. 2006;133:1457–1465. - PubMed
-
- Baehrecke EH. Autophagic programmed cell death in Drosophila. Cell Death Differ. 2003;10:940–945. - PubMed
-
- Beaulaton J, Lockshin RA. Ultrastructural study of the normal degeneration of the intersegmental muscles of Anthereae polyphemus and Manduca sexta (Insecta, Lepidoptera) with particular reference of cellular autophagy. J. Morphol. 1977;154:39–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
