

Transcriptional regulation of the citrate gene cluster of Enterococcus faecalis Involves the GntR family transcriptional activator CitO
- PMID: 18805984
- PMCID: PMC2576652
- DOI: 10.1128/JB.01704-07
Transcriptional regulation of the citrate gene cluster of Enterococcus faecalis Involves the GntR family transcriptional activator CitO
Abstract
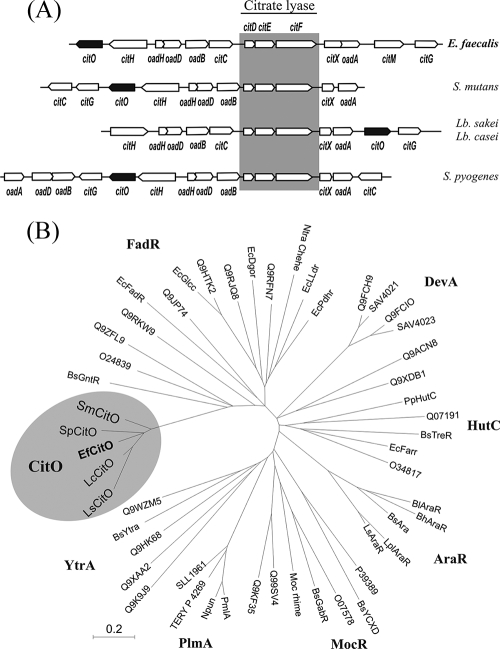
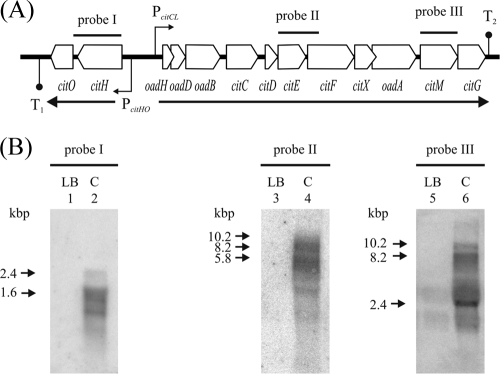
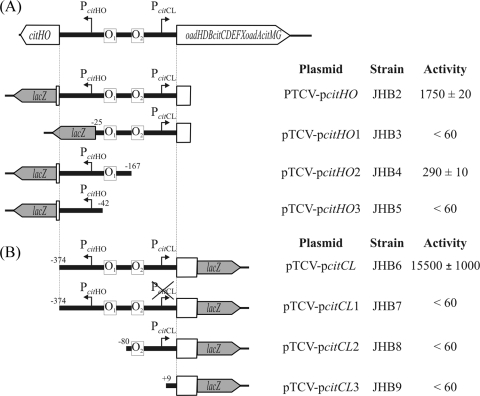
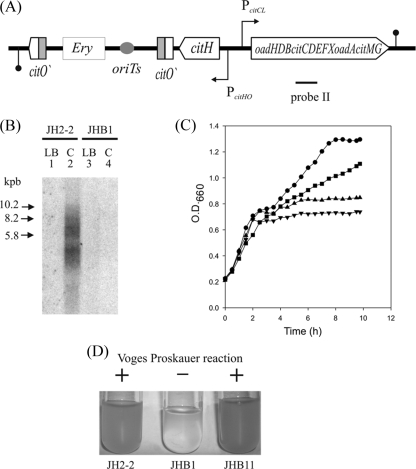
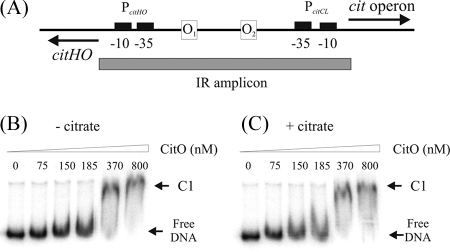
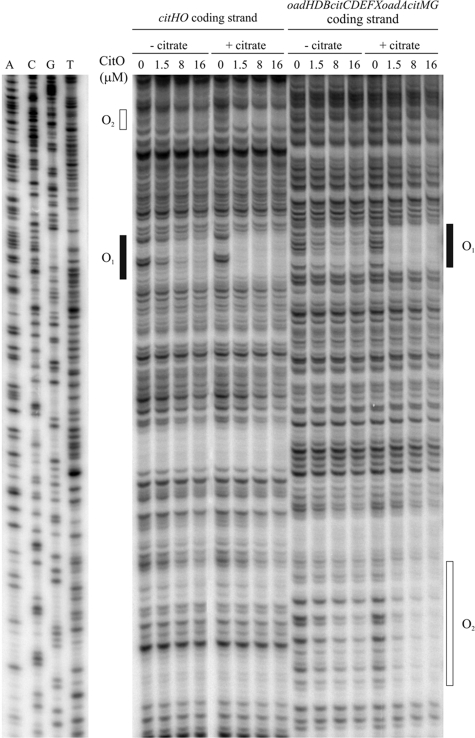
The genome of the gram-positive bacterium Enterococcus faecalis contains the genes that encode the citrate lyase complex. This complex splits citrate into oxaloacetate and acetate and is involved in all the known anaerobic bacterial citrate fermentation pathways. Although citrate fermentation in E. faecalis has been investigated before, the regulation and transcriptional pattern of the cit locus has still not been fully explored. To fill this gap, in this paper we demonstrate that the GntR transcriptional regulator CitO is a novel positive regulator involved in the expression of the cit operons. The transcriptional analysis of the cit clusters revealed two divergent operons: citHO, which codes for the transporter (citH) and the regulatory protein (citO), and upstream from it and in the opposite direction the oadHDB-citCDEFX-oadA-citMG operon, which includes the citrate lyase subunits (citD, citE, and citF), the soluble oxaloacetate decarboxylase (citM), and also the genes encoding a putative oxaloacetate decarboxylase complex (oadB, oadA, oadD and oadH). This analysis also showed that both operons are specifically activated by the addition of citrate to the medium. In order to study the functional role of CitO, a mutant strain with an interrupted citO gene was constructed, causing a total loss of the ability to degrade citrate. Reintroduction of a functional copy of citO to the citO-deficient strain restored the response to citrate and the Cit(+) phenotype. Furthermore, we present evidence that CitO binds to the cis-acting sequences O(1) and O(2), located in the cit intergenic region, increasing its affinity for these binding sites when citrate is present and allowing the induction of both cit promoters.
Figures


Similar articles
-
CcpA represses the expression of the divergent cit operons of Enterococcus faecalis through multiple cre sites.BMC Microbiol. 2011 Oct 11;11:227. doi: 10.1186/1471-2180-11-227. BMC Microbiol. 2011. PMID: 21989394 Free PMC article.
-
Acid-inducible transcription of the operon encoding the citrate lyase complex of Lactococcus lactis Biovar diacetylactis CRL264.J Bacteriol. 2004 Sep;186(17):5649-60. doi: 10.1128/JB.186.17.5649-5660.2004. J Bacteriol. 2004. PMID: 15317769 Free PMC article.
-
Aroma compounds generation in citrate metabolism of Enterococcus faecium: Genetic characterization of type I citrate gene cluster.Int J Food Microbiol. 2016 Feb 2;218:27-37. doi: 10.1016/j.ijfoodmicro.2015.11.004. Epub 2015 Nov 14. Int J Food Microbiol. 2016. PMID: 26594791
-
Anaerobic citrate metabolism and its regulation in enterobacteria.Arch Microbiol. 1997 Feb-Mar;167(2-3):78-88. Arch Microbiol. 1997. PMID: 9133329 Review.
-
Genetic organization and expression of citrate permease in lactic acid bacteria.Genet Mol Res. 2004 Jun 30;3(2):273-81. Genet Mol Res. 2004. PMID: 15266398 Review.
Cited by
-
Genomic comparative analysis of the environmental Enterococcus mundtii against enterococcal representative species.BMC Genomics. 2014 Jun 18;15(1):489. doi: 10.1186/1471-2164-15-489. BMC Genomics. 2014. PMID: 24942651 Free PMC article.
-
Bacillus subtilis Nucleoid-Associated Protein YlxR Is Involved in Bimodal Expression of the Fructoselysine Utilization Operon (frlBONMD-yurJ) Promoter.Front Microbiol. 2020 Aug 21;11:2024. doi: 10.3389/fmicb.2020.02024. eCollection 2020. Front Microbiol. 2020. PMID: 32983026 Free PMC article.
-
Comparative genomic analysis of pathogenic and probiotic Enterococcus faecalis isolates, and their transcriptional responses to growth in human urine.PLoS One. 2010 Aug 31;5(8):e12489. doi: 10.1371/journal.pone.0012489. PLoS One. 2010. PMID: 20824220 Free PMC article.
-
Identification of Potential Citrate Metabolism Pathways in Carnobacterium maltaromaticum.Microorganisms. 2021 Oct 18;9(10):2169. doi: 10.3390/microorganisms9102169. Microorganisms. 2021. PMID: 34683489 Free PMC article.
-
Tyramine biosynthesis in Enterococcus durans is transcriptionally regulated by the extracellular pH and tyrosine concentration.Microb Biotechnol. 2009 Nov;2(6):625-33. doi: 10.1111/j.1751-7915.2009.00117.x. Epub 2009 May 21. Microb Biotechnol. 2009. PMID: 21255297 Free PMC article.
References
-
- Bandell, M., M. E. Lhotte, C. Marty-Teysset, A. Veyrat, H. Prevost, V. Dartois, C. Divies, W. N. Konings, and J. S. Lolkema. 1998. Mechanism of the citrate transporters in carbohydrate and citrate cometabolism in Lactococcus and Leuconostoc species. Appl. Environ. Microbiol. 641594-1600. - PMC - PubMed
-
- Belitsky, B. R. 2004. Bacillus subtilis GabR, a protein with DNA-binding and aminotransferase domains, is a PLP-dependent transcriptional regulator. J. Mol. Biol. 340655-664. - PubMed
-
- Blancato, V. S., C. Magni, and J. S. Lolkema. 2006. Functional characterization and Me2+ ion specificity of a Ca2+-citrate transporter from Enterococcus faecalis. FEBS J. 2735121-5130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
