

Jade-1 inhibits Wnt signalling by ubiquitylating beta-catenin and mediates Wnt pathway inhibition by pVHL
- PMID: 18806787
- PMCID: PMC2830866
- DOI: 10.1038/ncb1781
Jade-1 inhibits Wnt signalling by ubiquitylating beta-catenin and mediates Wnt pathway inhibition by pVHL
Abstract
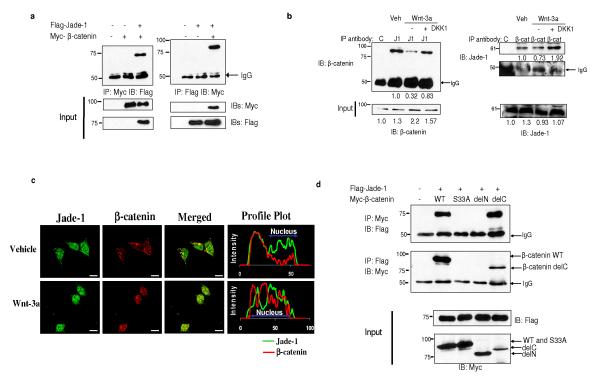
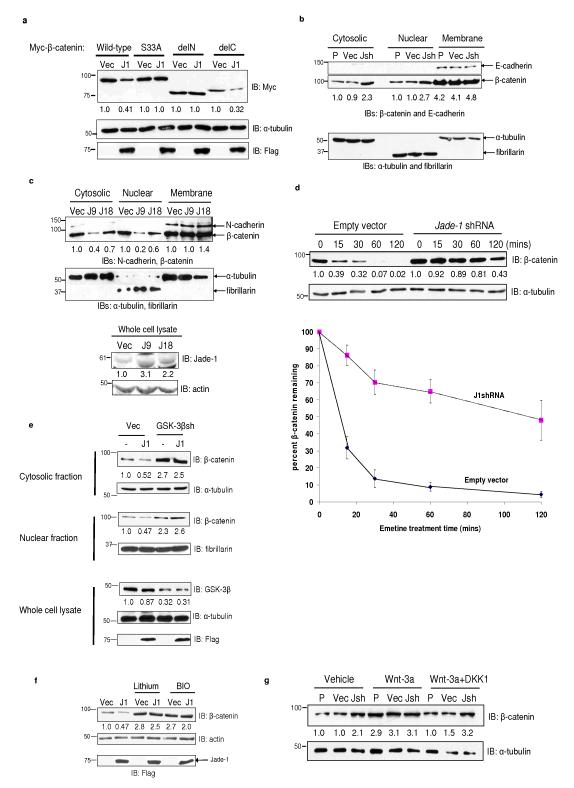
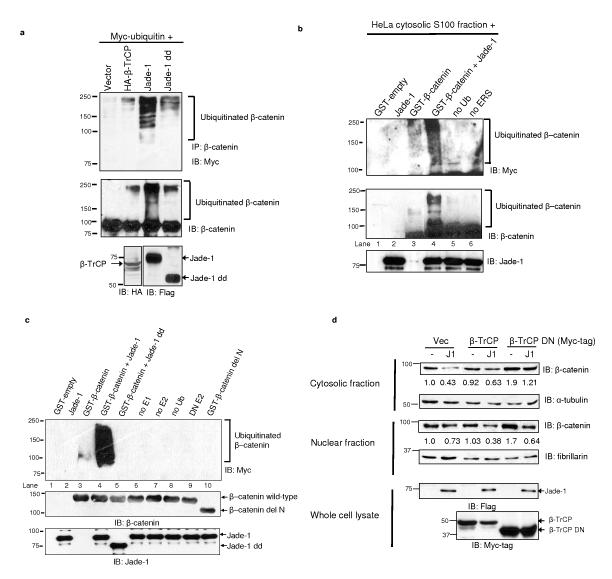
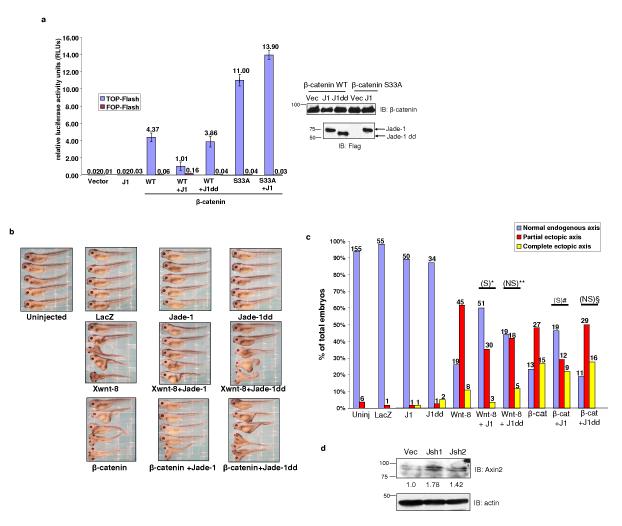
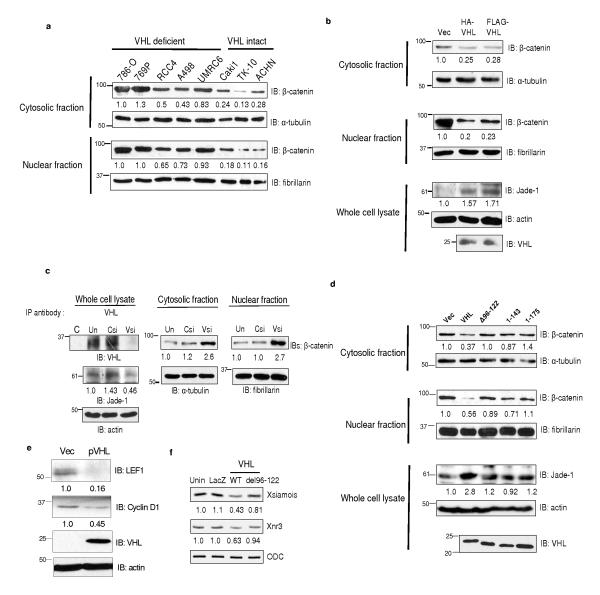
The von Hippel-Lindau protein pVHL suppresses renal tumorigenesis in part by promoting the degradation of hypoxia-inducible HIF-alpha transcription factors; additional mechanisms have been proposed. pVHL also stabilizes the plant homeodomain protein Jade-1, which is a candidate renal tumour suppressor that may correlate with renal cancer risk. Here we show that Jade-1 binds the oncoprotein beta-catenin in Wnt-responsive fashion. Moreover, Jade-1 destabilizes wild-type beta-catenin but not a cancer-causing form of beta-catenin. Whereas the well-established beta-catenin E3 ubiquitin ligase component beta-TrCP ubiquitylates only phosphorylated beta-catenin, Jade-1 ubiquitylates both phosphorylated and non-phosphorylated beta-catenin and therefore regulates canonical Wnt signalling in both Wnt-off and Wnt-on phases. Thus, the different characteristics of beta-TrCP and Jade-1 may ensure optimal Wnt pathway regulation. Furthermore, pVHL downregulates beta-catenin in a Jade-1-dependent manner and inhibits Wnt signalling, supporting a role for Jade-1 and Wnt signalling in renal tumorigenesis. The pVHL tumour suppressor and the Wnt tumorigenesis pathway are therefore directly linked through Jade-1.
Figures
Comment in
-
One hit, two outcomes for VHL-mediated tumorigenesis.Nat Cell Biol. 2008 Oct;10(10):1127-8. doi: 10.1038/ncb1008-1127. Nat Cell Biol. 2008. PMID: 18830218 No abstract available.
References
-
- Maxwell PH, et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399:271–5. - PubMed
-
- Cohen HT, McGovern FJ. Renal-cell carcinoma. N Engl J Med. 2005;353:2477–90. - PubMed
-
- Zhou MI, et al. The von Hippel-Lindau tumor suppressor stabilizes novel plant homeodomain protein Jade-1. J Biol Chem. 2002;277:39887–98. - PubMed
-
- Zhou MI, Wang H, Foy RL, Ross JJ, Cohen HT. Tumor suppressor von Hippel-Lindau (VHL) stabilization of Jade-1 protein occurs through plant homeodomains and is VHL mutation dependent. Cancer Res. 2004;64:1278–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
