

The energy hypothesis of sleep revisited
- PMID: 18809461
- PMCID: PMC2948963
- DOI: 10.1016/j.pneurobio.2008.08.003
The energy hypothesis of sleep revisited
Abstract
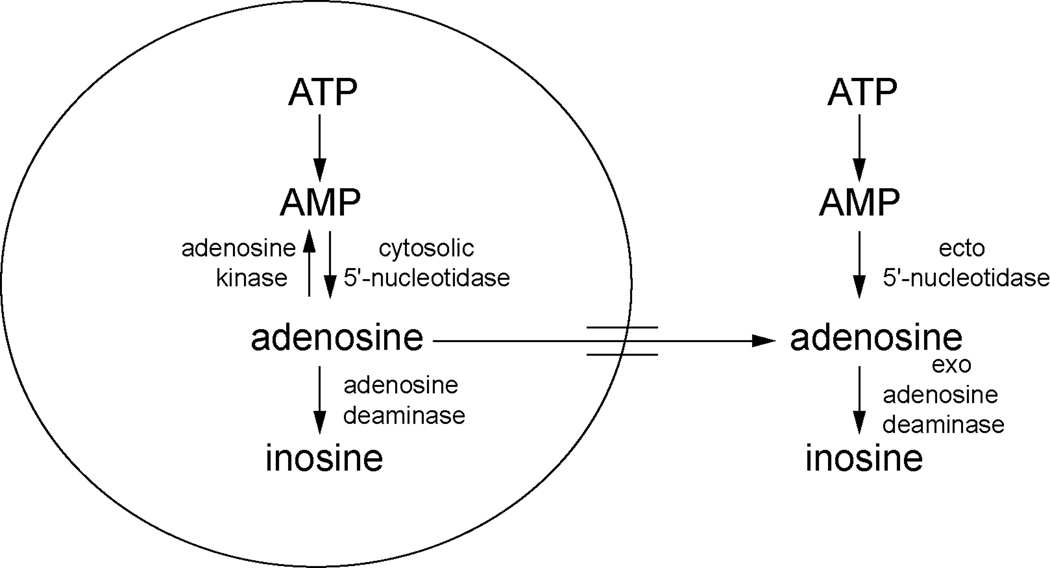
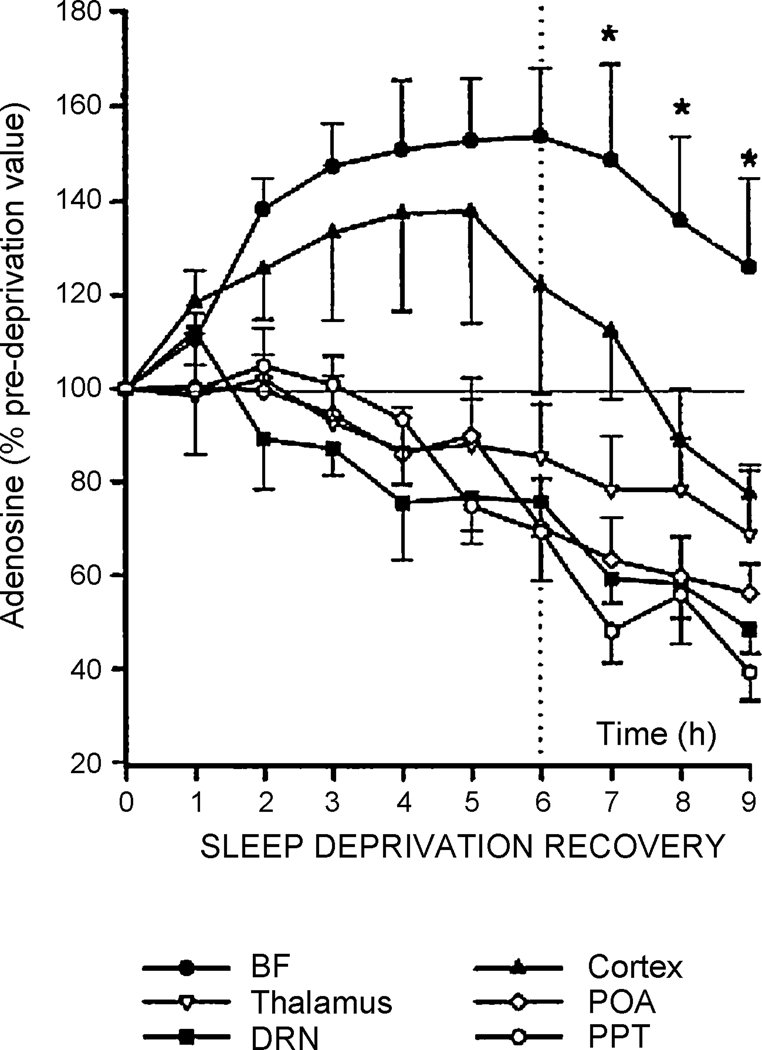
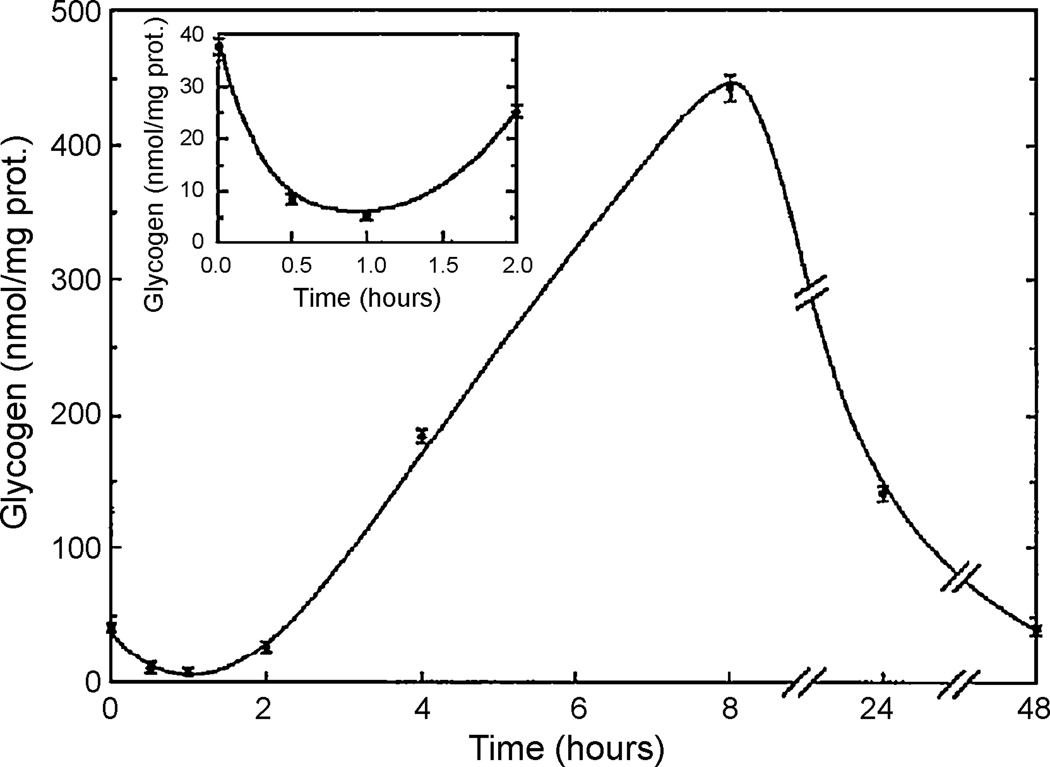
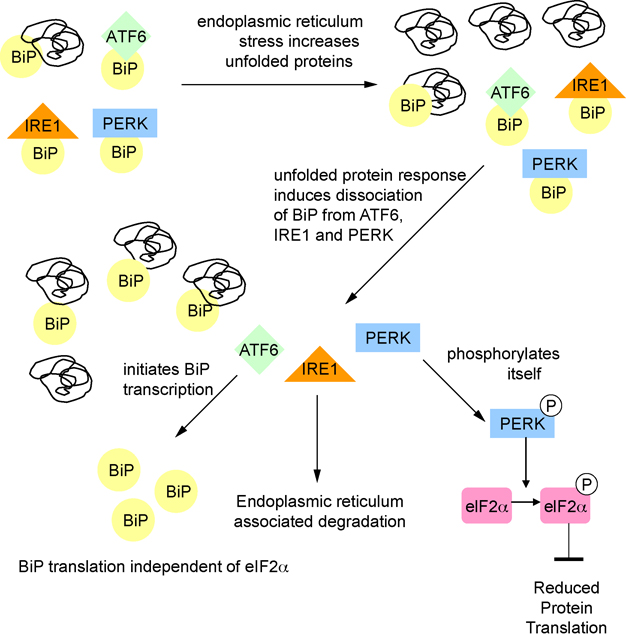
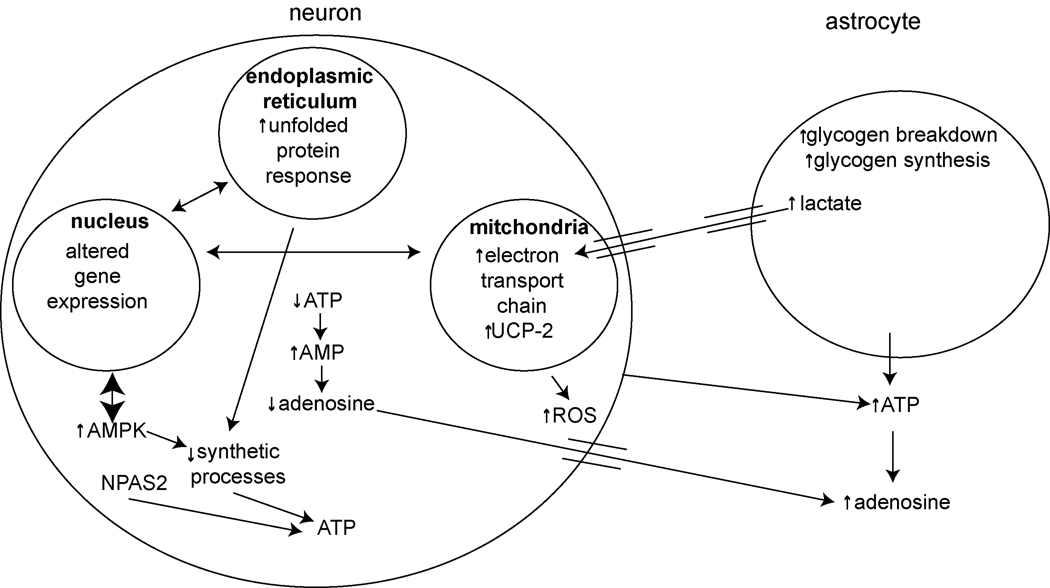
One of the proposed functions of sleep is to replenish energy stores in the brain that have been depleted during wakefulness. Benington and Heller formulated a version of the energy hypothesis of sleep in terms of the metabolites adenosine and glycogen. They postulated that during wakefulness, adenosine increases and astrocytic glycogen decreases reflecting the increased energetic demand of wakefulness. We review recent studies on adenosine and glycogen stimulated by this hypothesis. We also discuss other evidence that wakefulness is an energetic challenge to the brain including the unfolded protein response, the electron transport chain, NPAS2, AMP-activated protein kinase, the astrocyte-neuron lactate shuttle, production of reactive oxygen species and uncoupling proteins. We believe the available evidence supports the notion that wakefulness is an energetic challenge to the brain, and that sleep restores energy balance in the brain, although the mechanisms by which this is accomplished are considerably more complex than envisaged by Benington and Heller.
Figures
References
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. 2002
-
- Allaman I, Pellerin L, Magistretti PJ. Protein targeting to glycogen mRNA expression is stimulated by noradrenaline in mouse cortical astrocytes. Glia. 2000;30:382–391. - PubMed
-
- Allaman I, Pellerin L, Magistretti PJ. Glucocorticoids modulate neurotransmitter-induced glycogen metabolism in cultured cortical astrocytes. J Neurochem. 2004;88:900–908. - PubMed
-
- Andrews ZB, Diano S, Horvath TL. Mitochondrial uncoupling proteins in the CNS: in support of function and survival. Nat Rev Neurosci. 2005;6:829–840. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
