

Dominant membrane uncoupling by mutant adenine nucleotide translocase in mitochondrial diseases
- PMID: 18809618
- PMCID: PMC2638581
- DOI: 10.1093/hmg/ddn306
Dominant membrane uncoupling by mutant adenine nucleotide translocase in mitochondrial diseases
Abstract
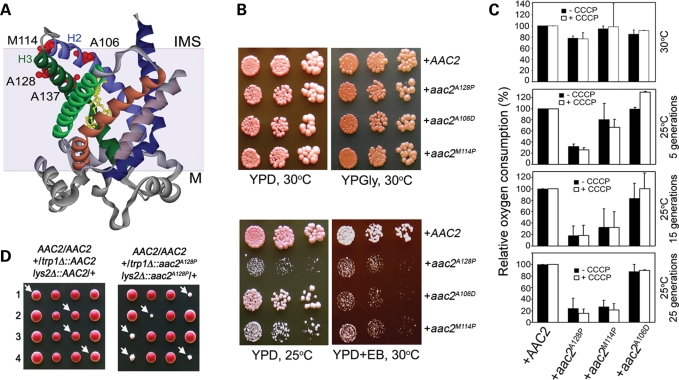
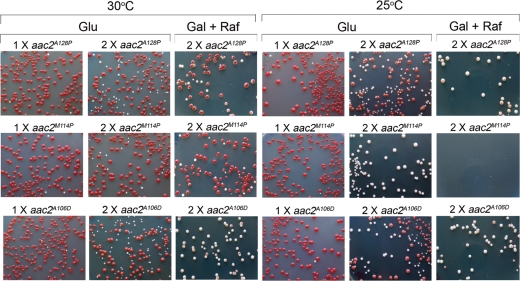
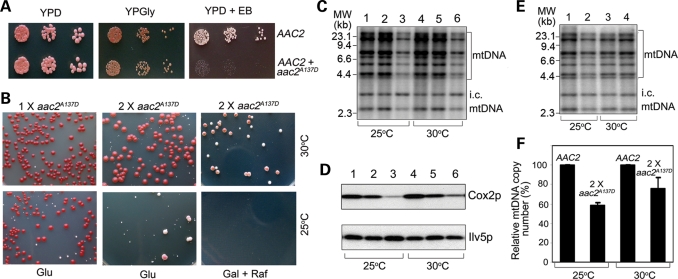
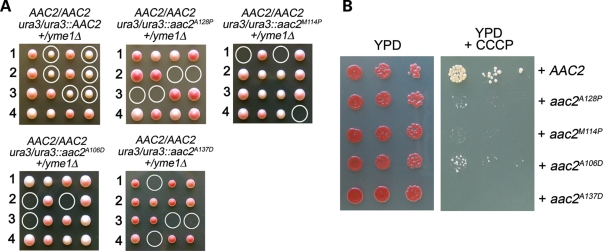
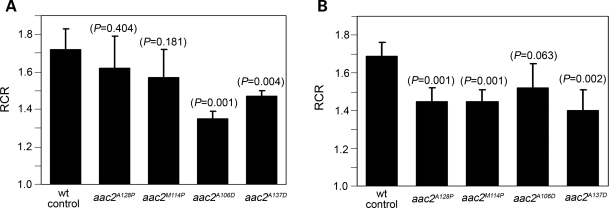
Adenine nucleotide translocase (Ant) is the most abundant protein on the mitochondrial inner membrane (MIM) primarily involved in ADP/ATP exchange. Ant also possesses a discrete membrane uncoupling activity. Specific mis-sense mutations in the human Ant1 cause autosomal dominant Progressive External Ophthalmoplegia (adPEO), mitochondrial myopathy and cardiomyopathy, which are commonly manifested by fractional mitochondrial DNA (mtDNA) deletions. It is currently thought that the pathogenic mutations alter substrate preference (e.g. ATP versus ADP) thereby dominantly disturbing adenine nucleotide homeostasis in mitochondria. This may interfere with mtDNA replication, consequently affecting mtDNA stability and oxidative phosphorylation. Here, we showed that the adPEO-type A128P, A106D and M114P mutations in the yeast Aac2p share the following common dominant phenotypes: electron transport chain damage, intolerance to moderate over-expression, synthetic lethality with low Deltapsi(m) conditions, hypersensitivity to the uncoupler carbonyl cyanide m-chlorophenylhydrazone (CCCP) and mtDNA instability. More interestingly, the aac2(A137D) allele mimicking ant1(A123D) in mitochondrial myopathy and cardiomyopathy exhibits similar dominant phenotypes. Because Aac2(A137D) is known to completely lack transport activity, it is strongly argued that the dominant mitochondrial damages are not caused by aberrant nucleotide transport. The four pathogenic mutations occur in a structurally dynamic gating region on the cytosolic side. We provided direct evidence that the mutant alleles uncouple mitochondrial respiration. The pathogenic mutations likely enhance the intrinsic proton-conducting activity of Ant, which excessively uncouples the MIM thereby affecting energy transduction and mitochondrial biogenesis. mtDNA disintegration is a phenotype co-lateral to mitochondrial damages. These findings provide mechanistic insights into the pathogenesis of the Ant1-induced diseases.
Figures
References
-
- Klingenberg M. The ADP and ATP transport in mitochondria and its carrier. Biochim. Biophys. Acta. 2008;1778:1978–2021. - PubMed
-
- Pebay-Peyroula E., Dahout-Gonzalez C., Kahn R., Trezeguet V., Lauquin G.J., Brandolin G. Structure of mitochondrial ADP/ATP carrier in complex with carboxyatractyloside. Nature. 2003;426:39–44. - PubMed
-
- Brustovetsky N., Klingenberg M. The reconstituted ADP/ATP carrier can mediate H+ transport by free fatty acids, which is further stimulated by mersalyl. J. Biol. Chem. 1994;269:27329–27336. - PubMed
-
- Kaukonen J., Juselius J.K., Tiranti V., Kyttala A., Zeviani M., Comi G.P., Keranen S., Peltonen L., Suomalainen A. Role of adenine nucleotide translocator 1 in mtDNA maintenance. Science. 2000;289:782–785. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
