

Selective expansion of a subset of exhausted CD8 T cells by alphaPD-L1 blockade
- PMID: 18809920
- PMCID: PMC2567485
- DOI: 10.1073/pnas.0801497105
Selective expansion of a subset of exhausted CD8 T cells by alphaPD-L1 blockade
Abstract
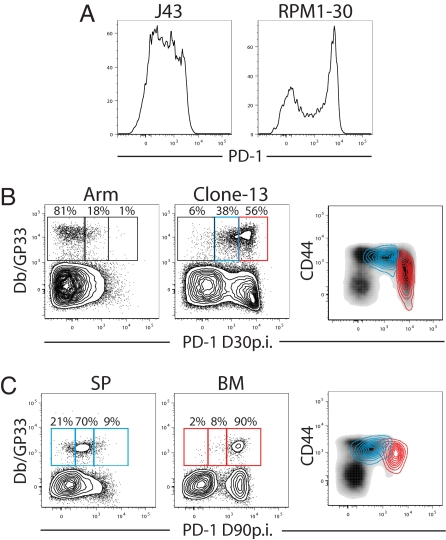
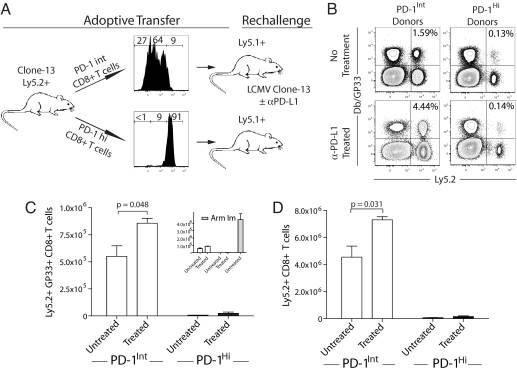
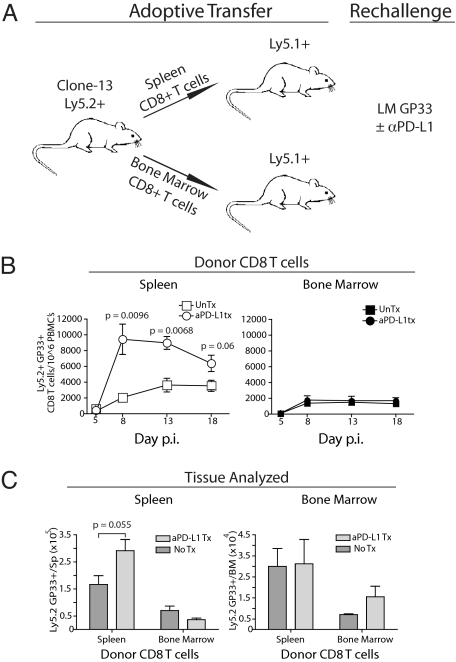
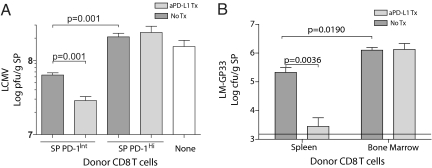
Programmed death-1 (PD-1) regulates T cell exhaustion during chronic infections. Blocking the PD-1:PD-ligand (PD-L) pathway reinvigorates exhausted CD8 T cells. Exactly how blocking PD-1:PD-L interactions improves T cell immunity, however, remains unclear. PD-1:PD-L blockade could reprogram all exhausted T cells to become antiviral effectors. Alternatively, this blockade might selectively expand a subset of exhausted T cells. We have identified two subpopulations of exhausted CD8 T cells during chronic viral infection in mice. One subset of exhausted CD8 T cells is rescued by alphaPD-L1 blockade, whereas the other subset appears more terminally differentiated and responds poorly to PD-1:PD-L blockade. Blocking PD-1:PD-L interactions reduces spontaneous apoptosis and enhances expansion and protective immunity of the rescuable subset, but not the more terminally differentiated subset of exhausted CD8 T cells. These results have implications for predicting clinical responses to PD-1-based therapeutic interventions and for understanding T cell dynamics during persisting infections.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol. 2007;8:239–245. - PubMed
-
- Barber DL, et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. - PubMed
-
- Isogawa M, Furuichi Y, Chisari FV. Oscillating CD8(+) T cell effector functions after antigen recognition in the liver. Immunity. 2005;23:53–63. - PubMed
-
- Day CL, et al. PD-1 expression on HIV-specific T cells is associated with T cell exhaustion and disease progression. Nature. 2006;443:350–354. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
